
Теплотехнический расчет
Результат| № п/п | Наименование расчётных параметров | Обозначения | Ед. измер. | Величина |
|---|---|---|---|---|
| 1 | Расчётная температура внутреннего воздуха | tв | °С | |
| 2 | Продолжительность отопительного периода | Zот.пер | сут | |
| 3 | Средняя температура наружного воздуха за отопительный период | tот.пер | °С | |
| 4 | Градусо/сутки отопительного периода | ГСОП | °С · сут |
| № п/п | Наименование расчётных параметров | Обозначения | Ед. измер. измер. | Величина |
|---|---|---|---|---|
| 1 | Коэффициент a | a | — | |
| 2 | Коэффициент b | b | — | |
| 3 | Требуемое сопротивление теплопередаче | Rтр | м2 · °С/Вт |
| № п/п | Наименование расчётных параметров | Ед. измер. | Величина | |
|---|---|---|---|---|
| 1 | Коэффициент теплоотдачи внутренней поверхности | α в | Вт/(м2 · С) | 8.7 |
| 2 | Коэффициент теплоотдачи наружной поверхности | α н | Вт/(м2 · С) |
Слои ограждающей конструкции
| № п/п | Наименование материала | ширина слоя, мм | Коэф. | Коэф. паропроницаеомсти, мг/(м·ч·Па) |
|---|
Страница не найдена
К вопросу о возможности получения двух урожаев картофеля для Московской обл.
O Ivashova, V Sychev, M Dyikanova, A Levshin, I Gasparyaz, Moscow Timiryazev Agricultural Academy,2020.
О ПРИЧИНАХ ПРОНИКНОВЕНИЯ ВИДОВ APATURA FABRICIUS, 1807 (LEPIDOPTERA, NYMPHALIDAE) В ЗАПАДНОЕ ЗАБАЙКАЛЬЕ
Гордеев С.Ю., Гордеева Т.В., Институт общей и экспериментальной биологии СО РАН, Улан-Удэ,2019.
Cовременная ситуация и ареал аскаридоза в России.
Н.А. Турбабина, Л.Ф. Морозова, ФГАОУ ВО Первый МГМУ им. И.М. Сеченова,2019.
Влияние способа получения исходных данных на прогнозные теплотехнические расчеты при проектировании в криолитозоне.
Г.П. Пустовойт, Э.С. Гречищева, С.И. Голубин, А.В. Аврамов, Московский государственный университет имени М.В. Ломоносова, 2018г.
Моделирование тепловых и гидравлических процессов магистральном газопроводе.
Базаров А.А., Данилушкин А.И., Самарский государственный технический университет, 2017
Изучение сортов овощной вигны при интродукции в Приморском крае.
журнал «Овощи России», 4 выпуск, 2016 г.
Зависимость заболеваемости острыми инфекциями дыхательных путей от суровости погоды в зимний период года (на примере города Кирова).
ГБОУ ВПО Кировская ГМА,2016г
Долгосрочное прогнозирование погоды посредством использования нейронных сетей.
VII Международная научно-практическая конференция «Приоритетные направления развития науки и образования», Якимчук А.
Поправочные коэффициенты на естественную продуктивность прудов водного тракта Кубань – Маныч.
Штефко Ю.Ю., Дементьев М.С. ФГАОУ ВПО «Северо-Кавказский федеральный университет», 2014г
Фундаментальные исследования. – 2014. – № 11-5. – С. 1094-1097;
Методика определения базового уровня энергопотребления и верификации измерений в рамках реализации энергосервисных контрактов.
Тупикина А.А. Новосибирский Государственный Технический Университет, г. Новосибирск
Риск простудных заболеваний горнорабочих.
Гудыма Н.Б.,Материалы VI Международной студенческой научной конференции «Студенческий научный форум», 2013г
Увы, закон энтропии работает 24 часа в сутки и уж не найти работ с 2007 по 2012гг. (:
(:
Изменение климатических данных и фактического значения ГСОП в Москве и их влияние на энергопотребление зданий
Одним из ключевых направлений повышения энергоэффективности экономики является снижение энергопотребления строящихся и эксплуатируемых зданий.
По данным [1], в Москве расход тепловой энергии на отопление многоквартирных домов (МКД) типовых серий по данным теплосчетчиков составляет 140-194 кВт•ч/м², что превышает современные нормативы энергопотребления.
Большой расход тепловой энергии связан с рядом факторов. Кроме конструкционных факторов есть и ряд других причин перерасхода тепла.
Для начала рассмотрим объективные причины, на которые повлиять нет возможности – погодные условия. Одной из причин перерасхода может являться различие фактических погодных условий эксплуатации от заложенных в проект.
В процессе проектирования при расчётах энергопотребления зданий учитываются климатические данные региона строительства.
ГСОП используются для нормирования сопротивления теплопередаче наружных ограждений зданий, сооружаемых в разных регионах страны и расчета удельного годового расхода тепловой энергии на отопление и вентиляцию.
Значение ГСОП численно равно произведению разности среднесуточной температуры наружного воздуха за отопительный период (ОП) tот и расчётной температуры внутреннего воздуха в здании tв на длительность ОП в сутках zот:
ГСОП = (tв– tот)•zот (формула 5.2, СП 50.13330.2012)
— где tот, — средняя температура наружного воздуха, °С, отопительного периода,
tв — расчётная температура внутреннего воздуха здания, °С.
Продолжительность отопительного периода для жилых и общественных зданий определяется из условия установления среднесуточной температуры наружного воздуха за 5-дневный период +8 ˚C, а для ряда медицинских и образовательных учреждений +10 ˚C.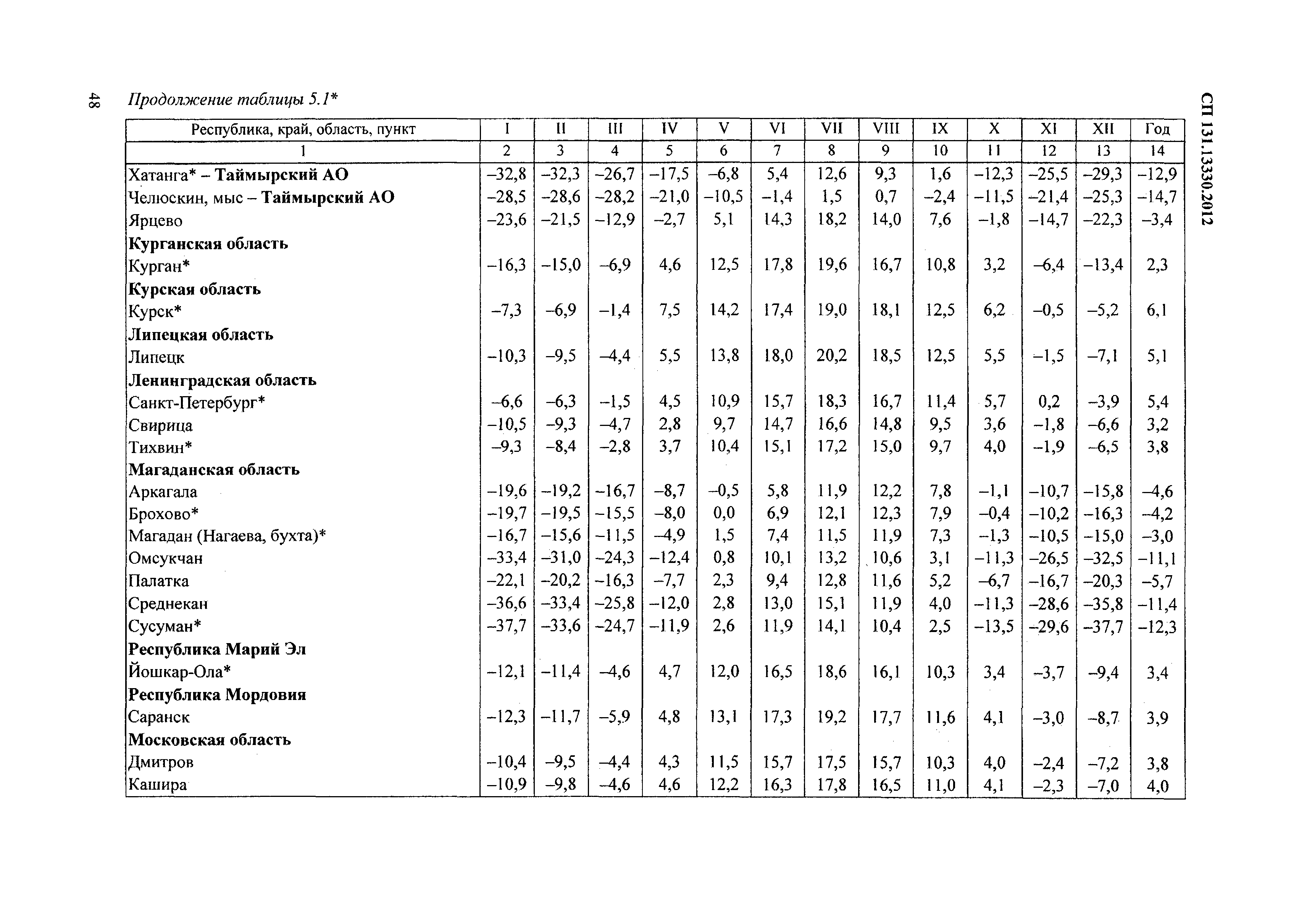
По многолетней практике эксплуатации большинства зданий при такой наружной температуре уровень внутренних тепловыделений, инсоляции и аккумулирующей способности здания не позволяют снижаться температуре внутреннего воздуха в помещениях ниже +18…+20 ˚C.
Согласно актуализированной редакции свода правил по строительной климатологии СП 131.13330.2012 в Москве по сравнению с требованиями СНиП 23-01-99* потеплело, а продолжительность отопительного периода сократилась. Средняя температура наружного воздуха за отопительный период для жилых зданий tот в СНиП 23-01–99* принималась равной –3,1ºС, а в СП 131.13330.2012 стала равна –2,2ºС. Продолжительность отопительного периода уменьшилась с 214 суток (СНиП 23-01–99*) до 205 суток (СП 131.13330.2012). В последней редакции — СП 131.13330.2018 эти данные не пересматривались.
В результате изменений расчётных климатических параметров расчетное значение ГСОП для жилых зданий, проектируемых в Москве снизилось с 4 943ºСсут (СНиП 23-01–99*), до 4 551ºСсут (СП 131.
На основе анализа климатических данных метеостанций, а также реального функционирования системы отопления г. Москвы за отопительные сезоны с 2005 по 2020 год, полученные из открытых источников были рассчитаны фактические значения ГСОП, представленные на
Рисунке №1 и в Таблице №1.
Рис.1 Диаграмма ГСОП по отопительным сезонам
Таблица №1.
В графическом виде распределение градусо-суток по месяцам за последние 6 сезонов представлено на Рис. № 2.
Рис.№ 2 Распределение градусо-суток по месяцам отопительных периодов.
Данные графики показывают, что максимальные значения градусо-суток в зависимости от сезона могут колебаться по разным месяцам (в сезонах с 2014 по 2020 — с декабря по март). При этом пропорционально градусо-суткам должно быть и потребление тепловой энергии зданиями при правильно организованной системе с погодным регулированием.
Данные фактических значений ГСОП для г. Москвы за отопительные сезоны с 2005 по 2020г демонстрируют, что для 11 отопительных сезонов фактическое значение ГСОП ниже требований СП по теплозащите, а в четырех сезонах выходит за рамки проектных значений, установленных в СП 131. 13330.2018 и СП 131.13330.2012. При использовании данных ГСОП из более ранней версии данных СП — СНиП 23-01–99* все отопительные сезоны укладываются в проектные рамки.
13330.2018 и СП 131.13330.2012. При использовании данных ГСОП из более ранней версии данных СП — СНиП 23-01–99* все отопительные сезоны укладываются в проектные рамки.
Колебания значений ГСОП по рассмотренным сезонам составляет 25%. Среднее значение ГСОП за рассмотренные сезоны составляет 4 293 ºС сут, что ниже проектного по данным актуальной версии СП 131.13330.2018.
Таким образом, фактические погодные условия в отдельные отопительные периоды могут быть более жёсткими, чем предусмотрено СП 131.13330.2018 и требовать большего удельного расхода тепла.
В целом фактическое значение ГСОП, рассчитанное по СП 131.13330.2018, соответствует проектным значениям и не может оказывать значительного влияния на наблюдаемое постоянное превышение фактического удельного расхода тепла зданиями во всех сезонах.
Следовательно, перерасход тепловой энергии обусловлен другими факторами.
При дальнейшем анализе энергопотребления домов в г. Москве были получены данные фактического потребления ряда зданий за период 2014-2018 годов и проведено их сопоставление с реальными погодными условиями данного сезона.
В большинстве случаев получились прямые зависимости расхода тепла от погодных условий, но в ряде случаев наблюдалось повышенное потребление тепла вне зависимости от климатических условий.
Для наглядного сопоставления приведем пример полученных данных.
Для примера данного превышения взяты два идентичных новых типовых панельных здания — 5-ти секционные 17-ти этажные дома серии П44Т в г. Москве. Типовые конструкции ограждающих конструкций, практически идентичная планировка и площади должны были привести к одинаковому расходу тепла зданий, но фактически расход тепла отличался более чем в 1,5 раза.
Диаграммы фактического потребления тепловой энергии домов приведены на Рисунке № 2, и Рисунке № 3.
Рис.№ 2. Удельный расход тепла: по проекту и по отопительным сезонам дома 1.
Рис. №3. Удельный расход тепла: по проекту и по отопительным сезонам дома 2.
Запуск системы отопления в доме 1 был произведен в 2014 году, в доме 2 в 2015 году. В доме 1 настроена система погодного регулирования. Первые сезоны после запуска отопления как правило производится отделка и постепенное заселение дома, поэтому данные сезоны непоказательны. Для окончательного анализа был принят отопительный сезон 2017/2018 года.
Первые сезоны после запуска отопления как правило производится отделка и постепенное заселение дома, поэтому данные сезоны непоказательны. Для окончательного анализа был принят отопительный сезон 2017/2018 года.
Удельное фактическое потребление тепловой энергии однотипных зданий различается в 1,69 раза. На двух панельных типовых 5-ти секционных 17-ти этажных домах серии П44Т при погодном регулировании удельное потребление энергии за сезон составило 151,1 Вт/м2 (рис. 2), а при отсутствии погодного регулирования 255,3 Вт/м2 (рис. 3).
Для большей наглядности составлены диаграммы фактического расхода тепла по месяцам (Рисунок № 4, Рисунок № 5).
Рис.№ 4. Сравнительная диаграмма удельного расхода тепловой энергии (Вт/м2) на отопление здания за сезон 2017-2018 г. дома 1.
Рис.№5. Сравнительная диаграмма удельного расхода тепловой энергии (Вт/м2) на отопление здания за сезон 2017-2018 г. дома 2.
Кривая расхода тепла у дома 1 повторяет с превышением кривую ГСОП данного сезона, а вот кривая фактического расхода тепловой энергии у дома 2 отличается от кривой ГСОП. Таким образом в доме 1 настроена система погодного регулирования, которая снижает и повышает расход тепла в зависимости от фактических погодных условий, а вот в доме 2 система погодного регулирования отсутствует – тепло подается постоянно без учета фактических погодных условий, что в итоге приводит к большому перерасходу тепловой энергии, а также отсутствию комфортных условий жильцов, так как для такого проживания при перерасходе тепловой энергии жильцам приходится прибегать к принудительному вентилированию, проветриванию помещений для снижения внутренней температуры помещений до приемлемых значений.
Таким образом в доме 1 настроена система погодного регулирования, которая снижает и повышает расход тепла в зависимости от фактических погодных условий, а вот в доме 2 система погодного регулирования отсутствует – тепло подается постоянно без учета фактических погодных условий, что в итоге приводит к большому перерасходу тепловой энергии, а также отсутствию комфортных условий жильцов, так как для такого проживания при перерасходе тепловой энергии жильцам приходится прибегать к принудительному вентилированию, проветриванию помещений для снижения внутренней температуры помещений до приемлемых значений.
Таким образом в качестве первоочередных мероприятий для снижения энергопотребления зданий должна применяться точная настройка системы отопления в соответствии с погодным регулированием, что позволит значительно снизить перерасход тепла и создание комфортных условий в здании.
__________________________________________________________________________________________________________
Список литературы:
- В.
 И. Ливчак. Градусо-сутки отопительного периода как инструмент сравнения уровня энергоэффективности зданий в России и в других странах. Энергосбережение №6’2015.
И. Ливчак. Градусо-сутки отопительного периода как инструмент сравнения уровня энергоэффективности зданий в России и в других странах. Энергосбережение №6’2015. - СП 131.13330.2018 Строительная климатология. Актуализированная редакция СП 131.13330.2012.
- СП 131.13330.2012 Строительная климатология. Актуализированная редакция СНиП 23-01-99* (с Изменениями N 1, 2).
- СНиП 23-01-99* Строительная климатология.
 И. Ливчак. Градусо-сутки отопительного периода как инструмент сравнения уровня энергоэффективности зданий в России и в других странах. Энергосбережение №6’2015.
И. Ливчак. Градусо-сутки отопительного периода как инструмент сравнения уровня энергоэффективности зданий в России и в других странах. Энергосбережение №6’2015.Статью подготовил инженер-эксперт
Отдела экспертиз зданий и сооружений на соответствие
теплотехническим и акустическим требованиям ГБУ «ЦЭИИС»
Бочков И.В.
| | Адрес этой страницы (вложенность) в справочнике dpva. ru: главная страница / / Техническая информация / / Физический справочник / / Климат. Климатические данные. Природные данные / / СНиП 23-01-99. Строительная климатология. (Статистика климатических данных) / / СНиП 23-01-99. Строительная климатология. Приложение А (справочное). Методы расчета климатических параметров ru: главная страница / / Техническая информация / / Физический справочник / / Климат. Климатические данные. Природные данные / / СНиП 23-01-99. Строительная климатология. (Статистика климатических данных) / / СНиП 23-01-99. Строительная климатология. Приложение А (справочное). Методы расчета климатических параметровПоделиться:
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Если Вы не обнаружили себя в списке поставщиков, заметили ошибку, или у Вас есть дополнительные численные данные для коллег по теме, сообщите , пожалуйста. Вложите в письмо ссылку на страницу с ошибкой, пожалуйста. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Коды баннеров проекта DPVA.ru Начинка: KJR Publisiers Консультации и техническая | Проект является некоммерческим. Информация, представленная на сайте, не является официальной и предоставлена только в целях ознакомления. Владельцы сайта www.dpva.ru не несут никакой ответственности за риски, связанные с использованием информации, полученной с этого интернет-ресурса.
Free xml sitemap generator Информация, представленная на сайте, не является официальной и предоставлена только в целях ознакомления. Владельцы сайта www.dpva.ru не несут никакой ответственности за риски, связанные с использованием информации, полученной с этого интернет-ресурса.
Free xml sitemap generator | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Использовалась сетчатка двойного экспоненциального распределения.
Использовалась сетчатка двойного экспоненциального распределения.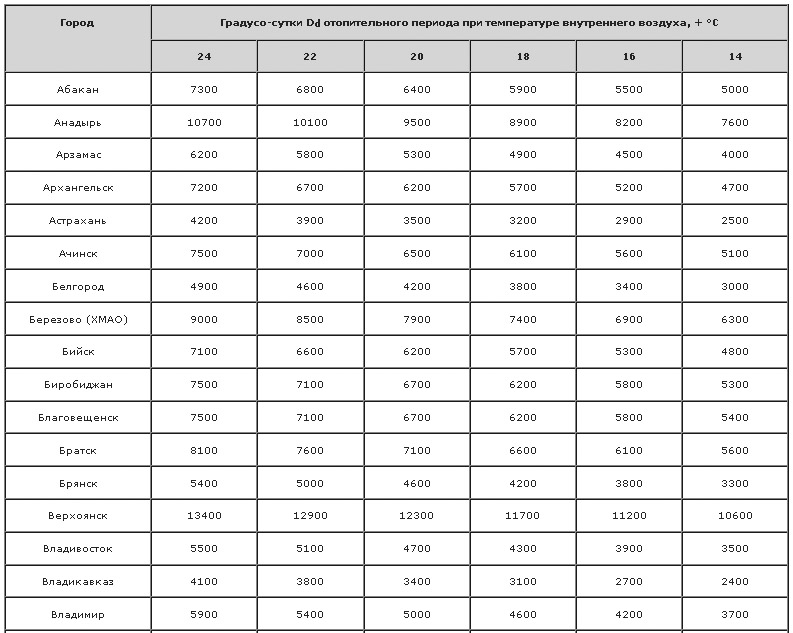


 таблицу А.2).
таблицу А.2).
(PDF) The Perm region zoning in terms of the heating period
2017 Географический вестник 4(43)
Метеорология
8. СНиП 23-01-99. Строительная климатология. М.: Стройиздат, 1999. 107 c.
9. СНиП 23-02-2003. Тепловая защита зданий. М.: Госстрой России, 2004. 25 с.
10. СП 131.13330.2012. Свод правил «Строительная климатология». Актуализированная редакция
СНиП 23-01-99. М.: Минстрой России, 2015.
11. Справочник эколого-климатических характеристик г. Москвы. Прикладные характеристики
климата, мониторинг загрязнения атмосферы, опасные явления, ожидаемые тенденции в XXI веке /
под ред. А.А. Исаева. М.: Изд-во Моск. ун-та, 2005. Т. 2. 411 с.
ун-та, 2005. Т. 2. 411 с.
12. Шкляев В.А., Шкляева Л.С. Характеристики отопительного периода в г. Перми и их временные
изменения // Географический вестник. 2011. №1(16). С. 44–48.
13. Энциклопедия климатических ресурсов Российской Федерации / под ред. Н.В.Кобышевой,
К.Ш. Хайруллина. СПб.: Гидрометеоиздат, 2005. 320 с.
14. ArcGISHelp. URL: http://resources.arcgis.com/ru/help/main/10.1/index.html (дата обращения:
2.07.2016). References
1. Anapolskaya, L.E. and Gandin, L.S. (1973), “Meteorologicheskie faktori teplovogo rezhima zdanii”,
[Meteorological factors of the thermal regime of buildings], Leningrad, Gidrometeoizdat, Russia.
2. Buligina O.N, Razuvaev, V.N. and Aleksandrova, T.M. (2014), “Opisanie massiva dannih sutochnoi
temperature vozduha i kolichestva osadkov na meteorologicheskih stantsiyah Rossii i bivshego SSSR (TTTR).
Svidetelstvo o gosudarstvennoi registratsii bazi dannih № 2014620942”, [Description data array of daily
temperature and precipitation at the meteorological stations of Russia and the former USSR (TTTR). The
The
certificate of state registration database № 2014620942], available at: http://meteo.ru/data/162-temperature-
precipitation#.
3. GOST R 55656-2013 (2014), “Energeticheskie harakteristiki zdanii. Raschet ispolzovaniya energii
dlya otopleniya pomeshchenii.(Proekt, pervaya redaktsiya)” [GOST R 55656-2013.The energy performance
of buildings.Calculation of energy use for space heating. (Draft first edition)], (ISO 13790:2008), Moscow,
Russia.
4. Zavarina, M.V. (1976), “Stroitelnaya klimatologiya”, [Building climatology], Gidrometeoizdat,
Leningrad, USSR.
5. Federal service for Hydrometeorology and environmental monitoring of Russia (2007), “Izmenenie
klimata: informatsionnyi bulluten (fevral – mart 2017)”, [Climate change: newsletter (February – March
2017)], no. 65, Moscow, Russia.
6. Kobisheva, N.V. (ed.) (2001), “Klimat Rossii”, [Climate of Russia], Gidrometeoizdat, St. Petersburg,
Russia.
7. Permyakov, J.A. and Gavrilyuk, G.D. (1985), “The heating season in Perm”, Voprosi mikro- i
mezoklimata, tsirkulyatsii i zagryazneniya atmosheri, pp. 78–84.
8. SNiP 23-01-99, (1999), Stroitelnaya klimatologiya. Sizmeneniyami 2003 g. [SNiP 23-01-99. Building
climatology.With the changes in 2003], Stroiizdat, Moscow, Russia.
9. Russian Federation State Committee for Construction and Housing and Municipal Economy (Gosstroy
Russia), (2004), SNiP 23-02-2003. Teplovaya zashchita zdanii. [SNiP 23-02-2003. Thermal protection of
buildings], Moscow, Russia.
10. The Ministry Of Construction Of Russia (2015), “SP 131.13330.2012: svod pravil “Stroitelnaya
klimatologiya” [SP 131.13330.2012, A set of rules «Construction climatology». Revised edition], Moscow,
Russia.
11. Isaev, A.A. (ed.) (2005), “Handbook of Moscow Environmental and Climatic Features. Vol. 2.
Applied Climatic Parameters, Air Pollution Monitoring, Dangerous Weather Phenomena, Expected
Tendencies at XXI Century”, Geography Department of Moscow State University Publishing.
12. Shklyaev,V.A. and Shklyaeva, L.S. (2011),“Harakteristiki otopitelnogo perioda v Permi i ih
vremennie izmeneniya”, [Characteristics of the heating period in the city of Perm and temporary changes],
GeographicalBulletin, no. 1 (16), pp. 44–48.
13. Kobisheva, N.V. and Hairullin, K.Sh. (ed.) (2005), “Encyclopedia klimaticheskih resursov Rossiiskoi
Federatsii” [Encyclopedia of climate resources of the Russian Federation], Gidrometeoizdat, St. Petersburg,
Russia.
14. ArcGIS Help, available at: http://resources.arcgis.com/ru/help/main/10.1/index.html (accessed
02.07.2016). Поступила в редакцию: 14.07.2017
Норматив температуры воздуха в квартире в отопительный сезон
В осенне-зимний период, когда температура опускается до 8 градусов и ниже, причем держится на этой отметке более пяти дней подряд, органы местного самоуправления объявляют начало отопительного сезона.
К сожалению, редкий поставщик услуг имеет четко отрегулированную систему теплоснабжения.
Когда температура в квартире мало отличается от температуры за окном либо наоборот, батареи раскалены и даже открытые форточки не особо спасают, не стоит сидеть, сложа руки. Руководствуясь СНиПом (Санитарные правила и нормы), обратитесь к коммунальщикам с претензией.
Вконтакте
Одноклассники
Мой мир
Норматив отопления
Для отстаивания собственных интересов и для того чтобы платить за качество, а не за цифры в квитанции, ознакомьтесь с существующими нормативами допустимой и оптимальной температуры в квартире на период отопительного сезона:
- 25 градусов в душевых и ванных комнатах, поскольку холод и высокая влажность негативно сказывается на здоровье человека. Однако допустимы и 18, и 26 градусов по крайним значениям нормы. Тот же регламент установлен и для комнат, объединяющих туалет и ванную;
- норма для жилых помещений – 20 — 22 градуса, допустим разброс от 18 до 24 градусов;
- в туалете и кухне нижняя граница нормы —18 градусов, верхняя — 26 градусов;
- в комнатах, расположенных по углам зданий – от 20 градусов;
- на лестничных клетках допускается температура от 14 до 20 градусов;
- подвалы и чердаки – 4 градуса тепла.
Важно: по желанию заказчика-застройщика лестничные марши могут не отапливаться, если в квартирах имеются системы теплоснабжения. То же относится и к районам с благоприятными климатическими условиями, где зимняя температура не опускается ниже пяти градусов. (СНиП 41-01-2003 «Отопление, вентиляция и кондиционирование»).
Семьи с детьми должны учитывать, что для здорового развития ребенка температура детских комнат должна составлять не менее 21 градуса, для новорожденных – от 24 градусов и выше. При необходимости, добрать градусы можно с помощью автономных обогревателей.
Но, если отопительный сезон наступил, а вы, приходя домой, снимаете шубу и надеваете тулуп и валенки – измерьте температуру в помещении.
[rek_custom1]
Как правильно делать замеры?
Прежде всего, запаситесь терпением и градусником. В каждой комнате с периодичностью в 60 минут следует измерять температуру на высоте в полутора метрах от пола и в метре от внешней стены комнаты. Обязательно составьте акт проведенных замеров в 2 экземплярах.
Следует учесть, что, согласно Постановлению Правительства №354, в ночное время отклонение может составлять до трех градусов от нормы, а в дневное время – с пяти часов утра до двенадцати ночи, запрещено понижение температуры ниже предельно допустимых значений.
Минимум 18 градусов для жилых и 20 для жилых угловых комнат, в остальных помещениях — согласно нормативам. Превышение максимально разрешенной температуры допускается не более чем на 4 градуса.
Совет: если произошла авария на теплотрассе, подача тепла в квартиры не должна прерываться более чем на 16 часов. В течение месяца общее количество часов без отопления не должно быть более 24.
В случае нарушения температурного режима стоит скооперироваться с соседями и сделать замеры по всему стояку отопления. Показатели нескольких квартир станут веским аргументом для перерасчета стоимости отопления.
Когда составленный акт попадет в руки представителей жилищно-коммунального хозяйства, будет назначена проверка качества. Коммунальщики сформируют комиссию и проведут замеры самостоятельно.
Поскольку не в их интересах делать перерасчет за услугу, не соответствующую нормативным актам, может оказаться, что в день прихода комиссии ваша квартира будет походить на тропический рай.
В этом случае и пригодится подписанный соседями акт. Не исключена и вероятность судебного урегулирования конфликта.
Обязательно проверьте сертификат прибора, которым комиссия будет делать замеры, его регистрацию и поверку. Погрешность прибора не должна превышать параметр в 0,1 градуса. Действия комиссии должны соответствовать «Методу контроля» ГОСТ 30494-96, пункт 4:
- замеры производят при наружной температуре воздуха не выше 5 градусов;
- при ясной погоде в светлое время суток измерения не проводятся;
- во время ручных замеров температура фиксируется не менее трех раз с интервалом в 5 минут;
- в автоматическом режиме показатели снимают в течение 2 часов;
- средняя величина замеров принимается за конечную температуру.
При фиксации поставки услуг ненадлежащего качества, на восстановление нормативного температурного режима отводится не более семи календарных дней.
Обратите внимание: в случае отклонения предельно допустимых параметров, перерасчет за отопление составляет 0,15% от стоимости услуги за каждый градус отклонения расчетного периода.
Может так случиться, что подача тепла в квартиру идет в соответствии с расчетными нормами, но комфортнее от этого не становится. В этом случае стоит обратить внимание на тепловой контур жилья – все, что, так или иначе, граничит с внешним периметром помещений.
Как добиться оптимальной температуры в квартире?
Прежде всего, убедитесь, что окна и двери надежно стыкуются со стенами. Даже небольшие щели способствуют выхолаживанию помещений. Если стыки негерметичны, утеплите любыми подручными средствами, включая вату и скотч-ленту.
Обладателям современных сплит-систем потребуется лишь задать определенный температурный параметр, и умная техника обеспечит комфортный режим.
Если же тепловой контур герметичен, а операторы котельной добросовестно исполняют свои обязанности, от духоты спасет своевременное проветривание. Так и градус понизится, и комнаты свежим воздухом наполнятся.
За неимением лучшего, можно использовать электрические обогреватели, либо газовые. Правда, догонять температуру до нормы придется за собственный счет, в дополнение к счету за отопление.
В продаже появились терморегуляторы, принцип действия которых схож с алгоритмом сплит-систем, но регулятор подключают к основному теплоносителю. Сторонние ресурсы, такие, как электроэнергия, не задействуются.
В любом случае, если с вас берут плату за оказанные услуги, ваше право и обязанность требовать их качественное предоставление. В настоящее время, когда граждан наделили правом выбора управляющей компании, добиться справедливости гораздо легче, нежели во времена централизованного обслуживания населения.
Не ленитесь фиксировать отклонения от допустимых параметров температурного режима. Доказать свою правоту сложно, но, руководствуясь вышеизложенным, можно.
О том, почему власти не успевают вовремя включить отопление, смотрите в следующем видео:
Вконтакте
Google+
Мой мир
Видите неточности, неполную или неверную информацию? Знаете, как сделать статью лучше?
Хотите предложить для публикации фотографии по теме?
Пожалуйста, помогите нам сделать сайт лучше! Оставьте сообщение и свои контакты в комментариях — мы свяжемся с Вами и вместе сделаем публикацию лучше!
Расчетная температура наружного воздуха для отопления
РАСЧЕТНЫЕ ПАРАМЕТРЫ НАРУЖНОГО И ВНУТРЕННЕГО ВОЗДУХА ДЛЯ РАСЧЕТА ОТОПЛЕНИЯ И ВЕНТИЛЯЦИИВ данном посте приведены основные положения и выдержки из действующих СНиП. Расчетная температура наружного воздуха для отопления — это и есть средняя температура холодной пятидневки, которая закладывается в расчет системы отопления частного дома. Это усредненная температура наиболее холодных пятидневок за восемь самых холодных зим из последних 50 лет. Такой подход позволяет, с одной стороны, быть готовыми к сильным морозам, которые случаются лишь раз в несколько лет, с другой — не вкладывать в проект излишних средств. В масштабах массовой застройки речь идет о весьма значительных суммах.
Расчетная температура наружного воздуха для отопления. Измерение температуры и относительной влажности наружного воздуха. Термогигрометр оконный ТГО-1
Знание основных климатических факторов и особенностей их влияния на эксплуатационные качества строительных материалов и конструкций позволят всем желающим спроектировать и построить дом своими силами, а также сделать его теплым, сухим и уютным. Все ограждающие конструкции дома предназначены для защиты помещения от атмосферных воздействий: холода, дождя, снега, ветра и пр., называются ограждающими. К ним относятся: наружные стены, окна, двери, крыша. Чтобы сделать дом теплым необходимо правильно выбрать материал, учитывая его теплозащитные свойства именно для ограждающих конструкций дома.
К физико-климатическим факторам района строительства относятся: температура и влажность, скорость и направление ветра, высота снежного покрова и количество выпадающих осадков, глубина промерзания грунта, количество солнечных и пасмурных дней в году. Какие же надо учитывать при строительстве теплого дома? Разумеется те, которые непосредственно влияют на изменение температуры и влажности конструкций здания и в той или иной мере определяют выбор материала и типа конструкций. Прежде всего, это расчетная температура наружного воздуха для отопления в районе строительства в холодный период года и величина градусо суток отопительного периода.
При определении теплозащитных качеств и выборе конструкций наружных ограждений принимают следующую расчетную температуру наружного воздуха: для легких ограждений – абсолютно минимальную температуру tа; для ограждений малой массивности – среднюю наиболее холодных суток tх.с; средней массивности – среднюю из средних температур наиболее холодных суток и наиболее холодной пятидневки; для массивных ограждений – среднюю наиболее холодной пятидневки tн. Для перекрытий над подвалами и подпольями принимают среднюю наиболее холодной пятидневки температуру tн, независимо от массивности ограждения.
Небольшое отступление от темы. Дорогие друзья, нижеприведенная ссылка приведет Вас на обучающий курс Зинаиды Лукьяновой Фотошоп с нуля в видеоформате 3.0. Курс содержит 82 урока, которые прекрасны по содержанию и понятны новичку. Здесь же приведены 5 бесплатных урока, просмотрев которые, я оформил заявку на полный курс и не жалею. Я рекомендую данный курс всем, кому не чуждо чувство прекрасного и кто хочет попробовать себя в удаленной работе по профессии Дизайнер. Приобретя данный курс, вы не будете вечерами ходить из угла в угол, вы не будете чесать пузо, лежа перед телевизором – вы будете работать, создавая прекрасное. И, как сказать, может это и станет вашим смыслом жизни. Я искренне желаю Вам удачи. Вот эта ссылка. Дерзайте! http://o.cscore.ru/28gig49/disc149
Различия между расчетными температурами наружного воздуха необходимо знать, чтобы правильно выбрать теплозащиту ограждения. Ведь потери тепла конструкцией в течение суток происходят неравномерно. В ночное время, когда воздух наиболее холодный, температура наружной поверхности стены снижается максимально, и постепенно стена начинает охлаждаться по толщине. Быстрота охлаждения конструкции зависит от ее способности усваивать и отдавать теплоту или от тепловой инерции. В бревенчатом срубе или в здании с массивными стенами в самый морозный день человек не ощущает холода. Но в том же помещении, если оно плохо отапливается, через несколько дней становится холодно, промозгло и неуютно: низкие температуры наружного воздуха вызвали резкое уменьшение температуры внутренней поверхности ограждающей конструкции. Поэтому остывший дом с массивными стенами приходится протапливать иногда и несколько дней.
В связи с этим для ограждающих конструкций большой инерционности расчетная температура наружного воздуха для отопления принимается равной средней температуре наиболее холодной пятидневки. Период в 5 суток принят потому, что его длительность достаточна для того, чтобы низкая температура наружного воздуха, установившаяся в течение этого периода, вызвала максимальное уменьшение температуры на внутренней поверхности стены. Для охлаждения ограждения малой инерционности достаточно одних суток, поэтому для их теплотехнического расчета принимается средняя температура наружного воздуха наиболее холодных суток.
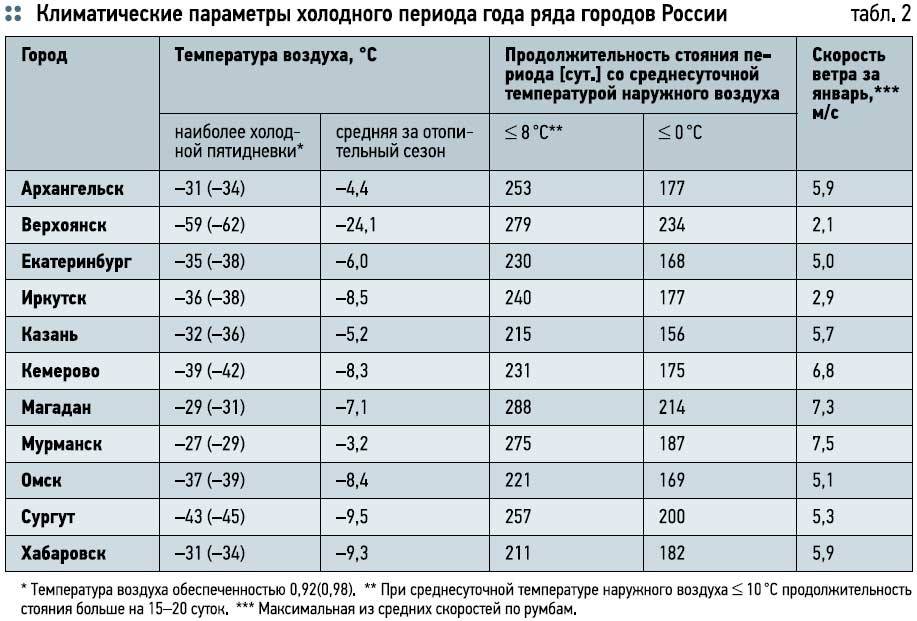
При проектировании системы отопления принимают такие расчетные температуры наружного воздуха: зимнюю, равную средней наиболее холодных пятидневок из восьми зим за 50-летний период tн и среднюю наиболее холодного периода (для вентиляции) tх.п. Расчетные параметры наружного воздуха для некоторых городов России (см. Таблицу 1).
РАСЧЕТНЫЕ ПАРАМЕТРЫ НАРУЖНОГО ВОЗДУХА ДЛЯ НЕКОТОРЫХ ГОРОДОВ РОССИИ (СНиП 2-А.6-72)Таблица 1
Город | Средняя температура наружного воздуха, оС | n, сут | v, м/с | Зона влажности | ||
| t н | t х.п | t о.п | ||||
| Архангельск | -32 | -19 | -4,7 | 251 | 5,9 | Влажная |
| Барабинск | -37 | -26 | -9,6 | 228 | 6,5 | Сухая |
| Барнаул | -39 | -23 | -8,3 | 219 | 5,9 | Сухая |
| Благовещенск | -34 | -25 | -11,5 | 212 | 3,4 | Нормальная |
| Верхоянск | -60 | -51 | -22 | 272 | 2,1 | Сухая |
| Владивосток | -25 | -16 | -4,8 | 201 | 9 | Влажная |
| Волгоград | -22 | -13 | -3.4 | 182 | — | Сухая |
| Екатеринбург | -31 | -20 | -6,4 | 228 | 5 | Сухая |
| Иркутск | -38 | -25 | -8,9 | 241 | 2,8 | Сухая |
| Калуга | -26 | -14 | -3,5 | 214 | 5 | Нормальная |
| Кемерово | -39 | -25 | -8,8 | 232 | 6,8 | Сухая |
| Красноярск | — 40 | -22 | — 7,2 | 235 | — | Сухая |
| Москва | -25 | -14 | -3,2 | 205 | 4,9 | Нормальная |
| Магадан | -35 | -23 | -9,6 | 278 | — | Нормальная |
| Новосибирск | -39 | -24 | -9,1 | 227 | 5,7 | Сухая |
| Омск | -37 | -23 | -7,7 | 220 | 5,1 | Сухая |
| Ростов | -22 | -8 | -0,6 | 175 | 6,5 | Сухая |
| С-Петербург | -25 | -11 | -2,2 | 219 | — | Влажная |
| Cахалин, г.Оха | -29 | -22 | -7,5 | 266 | 11,2 | Влажная |
| Томск | -40 | -25 | -8,8 | 234 | 3,3 | Нормальная |
| Тюмень | — 35 | -21 | — 5,7 | 220 | 3,9 | Сухая |
| Хабаровск | -32 | -23 | -10,1 | 205 | 5,9 | Влажная |
| Челябинск | -29 | -20 | -7,1 | 216 | 4,5 | Сухая |
| Чита | -38 | -30 | -11,6 | 240 | 3,9 | Сухая |
| Якутск | -55 | -45 | -19,5 | 254 | — | Сухая |
Что касается расчетной температуры внутреннего воздуха в доме, то она принимается в зависимости от назначения помещения (см. Таблицу 2). При расчетной температуре воздуха ниже t н = -31оС для угловой и прочих жилых комнат берутся более высокие значения, +22 и +20С (источник — постановление Правительства РФ от 23.05.2006 «Правила предоставления коммунальных услуг гражданам»).
РАСЧЕТНЫЕ ТЕМПЕРАТУРЫ ВНУТРЕННЕГО ВОЗДУХА В ПОМЕЩЕНИЯХ ЖИЛОГО ДОМА (СНиП 2-Л. 1-71)Таблица 2
| Наименование помещения | Температура внутреннего воздуха в помещении, оС |
| Жилая комната: угловая/прочие комнаты | 22/20 |
| Кухня | 18 |
| Коридор | 16 |
| Уборная | 16 |
| Ванная | 25 |
| Кладовая | 12 |
| Топочная | 14 |
t в– расчетная температура внутреннего воздуха отапливаемого помещения, оС.
ОПРЕДЕЛЕНИЕ ГРАДУСО СУТОК ОТОПИТЕЛЬНОГО ПЕРИОДА ДЛЯ г.ТОМСКАИ еще, мы должны определить градусо сутки отопительного периода (ГСОП). Формула для вычисления данного параметра имеет вид:
ГСОП=( t в — t о.п ) * n
Для г.Томска градусо сутки отопительного периода будут равны: ГСОП=( 20- (-8,8) * 234 = 6739,2 оС*сут. Для чего он используется и каким образом рассчитывается? От величины ГСОП будет зависеть нормируемое приведенное сопротивление теплопередаче ограждающих конструкций нашего дома. Например, для Московской области, где параметр ГСОП равен 4000 оС*сут, сопротивление теплопередаче ограждающей конструкции должно быть не меньше: для стен – 2,8 м2*оС /Вт, для перекрытий (пол 1-ого этажа, чердак или потолок мансарды) — 3,7 м2*оС /Вт, для окон и дверей – 0,35 м2*оС /Вт. В пункте №5 нашего плана по расчету отопления частного дома мы поговорим об этом поподробней. Приведенное сопротивление теплопередаче будет определено для всех ограждающих конструкций нашего дома.
Итак, расчетные параметры наружного и внутреннего воздуха для расчета отопления и вентиляции для нашего дома принимаем следующие:
t н = — 40оС — расчетная зимняя температура наружного воздуха для проектирования отопления;
t х.п = — 25 оС — расчетная зимняя температура наружного воздуха для проектирования вентиляции;
t о.п = — 8,8 оС — средняя температура отопительного периода;
n = 234 сут. — продолжительность отопительного периода;
v = 3,3 м/с — средняя скорость ветра за январь;
ГСОП= 6739,2 оС*сут.
Дорогие друзья, в следующем посте мы с вами произведем расчет тепловой нагрузки на отопление дома различными способами, сравним результаты и проанализируем их. Сегодня мы с вами выполнили 1-ый пункт нашего плана по расчету системы отопления дома — определили расчетную температуру наружного воздуха для отопления, а также определили градусо сутки отопительного периода для г. Томска. Кто еще не успел присоединяйтесь! До связи.
С уважением, Григорий Володин
Новые SNP в гене HSPB8 и их связь с признаками термостойкости у коренного рогатого скота сахивалов
Белки теплового шока (HSP) экспрессируются в ответ на тепловой стресс, а полиморфизм генов HSP на однонуклеотидном уровне, как сообщается, связан с термостойкостью и характеристиками продуктивности крупного рогатого скота. Ген HSPB8 был картирован на аутосоме 17 Bos taurus (BTA-17), охватывающей почти 13 252 п.н. и состоящей из трех экзонов и двух интронов.Настоящее исследование было проведено на коровах сахивал (n = 108), выращенных в субтропическом климате, с целью выявления SNP во всех трех экзонах и части интрона 1 гена HSPB8 и анализа их связи с признаками теплостойкости у коров сахивал. Частоту дыхания (ЧД) и ректальную температуру (RT) регистрировали один раз в вероятные экстремальные часы в разные сезоны или индекс температуры-влажности (THI), то есть зимой, весной и летом. Для проверки приспособляемости животных в период теплового стресса также рассчитывали коэффициент термостойкости (HTC).Сравнительный анализ последовательностей выявил всего два однонуклеотидных полиморфизма (SNP), то есть g.507G> A в экзоне 1 и g.881T> C в интроне 1 гена HSPB8. Из этих двух идентифицированных SNP только один SNP, то есть g.507G> A, оказался в значительной степени связанным с индикаторными признаками теплостойкости (RR, RT и HTC) у коров Сахивал. Изучение результатов в разные сезоны показало значительную (P <0,01) разницу в RR, RT и HTC между зимним, весенним и летним сезонами.Было обнаружено, что RR, RT и HTC были значительно ниже (P <0,01) в GA по сравнению с генотипом GG g.507G> SNP гена HSPB8. Однако в случае другого SNP, то есть g.881T> C, расположенного в интроне 1, RR, RT и HTC имели незначительную ассоциацию с разными генотипами, то есть TT и TC. Эти результаты могут частично предполагать, что генотип GA SNP g.507G> A гена HSPB8 имеет вероятную роль в термостойкости у сахивальского крупного рогатого скота и, следовательно, может использоваться в качестве маркера при выращивании термоустойчивого крупного рогатого скота в жарком тропическом и субтропическом климате.Тем не менее, нельзя игнорировать участие других регуляторных механизмов.
Ключевые слова: HSP; HSPB8; SNP.
Общегеномное ассоциативное картирование для идентификации локусов количественных признаков для ректальной температуры во время теплового стресса у крупного рогатого скота голштинской породы
Идентификация QTL, связанного с RT
Wang et al. [19] в ходе моделирования обнаружили, что корреляция QTL с суммой соседних SNP увеличивалась до 8 SNP, а затем уменьшалась по мере увеличения количества суммированных SNP.Это связано с тем, что ближайший к QTL SNP не всегда является лучшим предиктором эффекта QTL [25]. В текущем исследовании 2, 3, 4, 5 и 10 соседних SNP были усреднены для выявления консенсусных геномных областей, связанных с RT. GWAS, основанный на ssGBLUP, использовался вместо более традиционных подходов GWAS, поскольку последние не используют напрямую фенотипы негенотипированных людей. P-значения не использовались для объявления регионов значимыми, потому что такие значения трудно определить и сравнить с использованием классических тестов гипотез, когда используются уменьшенные оценки, такие как PTA [26].Недавние исследования также показали, что традиционные методы GWAS часто дают большое количество (например, сотни) значительных эффектов, большинство из которых не могут быть подтверждены в последующих исследованиях [27]. Основная цель этого исследования состояла в том, чтобы идентифицировать области генома, связанные с регуляцией RT, и это не зависит от тестов статистической значимости, которые более подходят для исследований на основе генов-кандидатов.
Доля дисперсии SNP, объясняемая каждым маркером в отдельности, показана на рисунке S1 и в файле S1.Отдельные результаты SNP были очень зашумленными, и только одна область на BTA 26 показала четкий сигнал. Использование окон 2-SNP (рис. S2) привело к более плавным эффектам маркеров, но по-прежнему было трудно отличить маркеры с большими эффектами от маркеров с небольшими эффектами. Графики Манхэттена из окон 3 и 5 SNP (рисунки 1 и 2) привели к более четким и в целом последовательным схемам. Результаты были аналогичными для окон 4-SNP (рисунок S3).
Результаты 10-SNP (рисунок S4) аналогичны результатам, полученным с использованием более узких окон.Тем не менее, анализ 10-SNP создает проблемы, потому что Illumina BovineSNP50 BeadChip имеет средний интервал между маркерами 49,4 кбит / с, так что каждая точка на результирующем графике Манхэттена покрывает в среднем 494 кбит / с. Это проблематично, потому что полезное равновесие сцепления (LD) у коровы простирается менее чем на 100 т.п.н. [28]. Гены или группы генов, которые не находятся в LD друг с другом, но которые имеют большие эффекты, должны быть представлены отдельными пиками.
20 крупнейших объясняющих локусов для RT для каждого из анализов перечислены в таблицах 2 и 3 и с S1 по S4.Чаще всего наибольшая доля дисперсии объясняется маркерами, фланкирующими область между 28 877 547 и 28 907 154 п.н. на аутосоме Bos taurus (BTA) 24. Эта область фланкируется сплайсосомной РНК U1 ( U1 ) слева (28 822 883). до 28,823,043 п.н.) и ген кадгерина-2 ( NCAD ) справа (от 28,992,666 до 29,241,119 п.н.). Малый рибонуклеопротеин U1 участвует в посттранскрипционной модификации и регуляции длины мРНК [29], оба из которых могут быть связаны с изменениями экспрессии генов в клетках, подвергнутых воздействию повышенной температуры [30], [31]. NCAD был более высоко экспрессирован у зараженного паратуберкулезом крупного рогатого скота, чем у неинфицированных животных [32], но неясно, какая связь, если таковая имеется, существует между NCAD и реакциями на стресс.
Две интересующие области были отмечены на BTA 16. Анализ 2- и 3-SNP (таблица S2 и 2) показал, что SNP на 58 500 249 п.н. объясняет 0,08 и 0,11% дисперсии SNP, соответственно. Этот контиг включает небольшую ядрышковую РНК ( SNORA19 ) от 58 520 021 до 58 520 149 п.н .; убиквитин-протеин-лигаза ( RFWD2 ) от 58 600 678 до 58 838 844 п.н .; и специфическая для малых тельцов Кахаля РНК 3 ( SCARNA3 ) с 58 628 128 до 58 628 268 п.н.Небольшие ядрышковые РНК, такие как SNORA19 , идентифицируют сайты образования псевдоуридина [33], которые могут участвовать в инициации трансляции. Ген SCARNA3 также кодирует небольшую ядрышковую РНК, сходную по структуре с SNORA19 . Ген RFWD12 кодирует убиквитин-протеинлигазу E3 RING-типа, которая отбирает белки для протеасомной деградации [34]. Многие аномальные белки протеолизируются системой убиквитина как часть стрессовой реакции у эукариот [35].Более того, всего 28 генов, идентифицированных как регулируемые тепловым шоком у эмбриона крупного рогатого скота, связаны с убиквитином С [31].
Второй интересующий регион БТА 16 расположен на отметке 35 272 426 б.п. Ближайшие гены к этой области — центросомный белок 170 кДа ( CEP170 ) и неактивная фосфолипаза D5 ( PLD5 ). Ни один из генов не имеет очевидной связи с физиологической реакцией на тепловой стресс.
Область BTA 5 приблизительно 89 500 000 п.н. была последовательно идентифицирована во время анализов.Два гена фланкируют консенсусную область, член семейства переносчиков органических анионов растворенного вещества 1C1 ( SLCO1C1 ) и фосфодиэстеразу ( PDE3A ). У человека SLCO1C1 опосредует Na + -независимый высокоаффинный транспорт тироксина и обратного трийодтиронина [36]. Возможно, этот ген участвует в механизме, который снижает концентрацию тироксина в плазме у молочных коров, подвергшихся тепловому стрессу [37]. Было показано, что человеческий белок теплового шока 70 увеличивает ферментативную активность фосфодиэстеразы в клетках, подвергшихся тепловому шоку [38], и может быть аналогичная ассоциация у крупного рогатого скота.
Анализ 4- и 5-SNP показал пик около 64 400 000 п.н. на BTA 4, что объясняет 0,09–0,11% наблюдаемой дисперсии (таблицы S3 и 3). Эта область включает гены келча-повтора и белка, содержащего домен BTB ( KBTBD2 ), и U6 snRNA-ассоциированного Sm-подобного белка LSM5 ( LSM5 ). KBTBD2 кодирует белок, который участвует в убиквитинировании белков [39], тогда как LSM5 , вероятно, участвует в процессинге РНК и формирует часть стрессовых гранул, наблюдаемых в стрессовых клетках, которые содержат мРНК, остановленные в трансляции [40].
Область с центром на SNP на 20 290 497 п.н. на BTA 26 объясняет от 0,10% до 0,22% наблюдаемой дисперсии в анализах 2-SNP-5-SNP (таблицы 2 и 3 и S1 – S3. SNP находится в глутамине). -оксалоуксусная трансаминаза, растворимый ген ( GOT1 ). GOT1 участвует в синтезе диоксида серы, который, как было показано, снижает образование реактивных продуктов кислорода и защищает ткань миокарда крысы в ответ на изопротеренол [41].
В более раннем исследовании Hayes et al.[42] идентифицировали SNP на BTA 29 (BFGL-NGS-30169), который был связан с генетической изменчивостью в эффектах теплового стресса на удои у голштинов и трикотажных животных. Соседний SNP на BTA 29 (ARS-BFGL-NGS-107395 на 47 527 067 пар оснований) был одним из 30 лучших маркеров в анализах SNP 2–5, занимая место между 19 и 35 наибольшими SNP. Он расположен на 355 843 п.н. выше SNP, идентифицированного Hayes et al. [42] в контиге, который включает только аннотированную небольшую ядрышковую РНК ( SNORD14 ).
Существуют небольшие, но значимые генетические корреляции RT во время теплового стресса с 305-дневными надоями молока, жира и белка, продуктивной жизнью, чистыми заслугами, показателем соматических клеток и частотой беременностей дочерей [8]. Независимо от анализа, использованного для определения SNP, объясняющих RT, ни один из 10 крупнейших объясняющих локусов для RT не был общим с 1586 маркерами SNP, относящимися к 31 другому признаку молочного скота, ранее идентифицированному Cole et al. [11]. Таким образом, можно было бы использовать SNP, связанные с RT, для отбора по термотолерантности без случайного выбора по другим признакам.
В заключение, эти данные демонстрируют, что QTL существует для предсказания некоторых генетических вариаций в RT. Эти QTL могут оказаться важными для генетического отбора термотолерантности. Проблема выполнения GWAS для признаков с низкой наследуемостью при низком количестве фенотипированных животных означает, что подтверждение полезности этих QTL имеет важное значение. Кроме того, было идентифицировано несколько генов-кандидатов для регуляции RT у молочного скота, и один или несколько из них могут играть важную роль в физиологической адаптации к тепловому стрессу.Только один ген-кандидат, SLC01C1 , который участвует в регуляции скорости метаболизма посредством транспорта тироксина, играет известную роль в процессах, контролирующих температуру тела. Чаще всего гены-кандидаты играют важную роль для стабилизации клеточной функции во время стресса. Среди них GOT1 , который синтезирует циторотективное соединение диоксид серы, гены, участвующие в убиквитинизации белков ( KBTBD2 и RFWD12 ), и гены, участвующие в метаболизме РНК ( LSM5, SCARNA3, SNORA19, и U1 ).
Определение однонуклеотидного полиморфизма (SNP) на основе гранул с использованием температуры плавления на микрочипе
Apple FS, Christenson RH, Valdes R, Andriak AJ, Berg A, Duh SH, Feng YJ, Jortani SA, Johnson NA, Koplen B , Mascotti K, Wu AHB (1999) Одновременное быстрое измерение миоглобина цельной крови, креатинкиназы MB и сердечного тропонина I с помощью сортировочной кардиологической панели для выявления инфаркта миокарда. Clin Chem 45: 199–205
Google Scholar
Ботштейн Д., Уайт Р.Л., Сколник М., Дэвис Р.В. (1980) Создание карты генетического сцепления у человека с использованием полиморфизмов длины рестрикционных фрагментов.Am J Hum Genet 32: 314–331
Google Scholar
Chen Y-H (2008) Идентификация однонуклеотидных полиморфизмов, связанных с размером помета, из дифференциально регулируемых генов ранних эмбрионов у свиноматок породы Ландрас. Магистерская диссертация, Национальный Тайваньский университет
Ди Карло Д., Агдам Н., Ли Л.П. (2006) Концентрации одноклеточных ферментов, кинетика и анализ ингибирования с использованием массивов гидродинамической изоляции клеток высокой плотности.Anal Chem 78: 4925–4930
Статья Google Scholar
Дрессман Д., Ян Х., Траверсо Г., Кинзлер К.В., Фогельштейн Б. (2003) Преобразование отдельных молекул ДНК во флуоресцентные магнитные частицы для обнаружения и подсчета генетических вариаций. Proc Natl Acad Sci 100: 8817–8822
Статья Google Scholar
Эддингс М.А., Джонсон М.А., Гейл Б.К. (2008) Определение оптимальной техники связывания ПДМС – ПДМС для микрофлюидных устройств.J Micromech Microeng 18: 067001
Артикул Google Scholar
Elenitoba-Johnson KSJ, Bohling SD (2001) Сканирование на основе раствора для одноосновных изменений с использованием красителя, связывающего двухцепочечную ДНК, и профилей флуоресценции-плавления. Am J Pathol 159: 845–853
Статья Google Scholar
Gundry CN, Vandersteen JG, Reed GH, Pryor RJ, Chen J, Wittwer CT (2003) Анализ плавления ампликона с мечеными праймерами: метод с закрытой пробиркой для дифференциации гомозигот и гетерозигот.Clin Chem 49: 396–406
Статья Google Scholar
Han C, Zhang Q, Ma R, Xie L, Qiu T, Wang L, Mitchelson K, Wang J, Huang G, Qiao J, Cheng J (2010) Интеграция улавливания одиночных ооцитов, экстракорпорального оплодотворения и эмбриона культивирование в микрожидкостном устройстве с микропланшетной структурой. Lab Chip 10: 2848–2854
Артикул Google Scholar
Hannewijk J, Haighton AJ (1958) Дифференциальный термический анализ.3. Кривые плавления масел и жиров. J Am Oil Chem Soc 35: 457–461
Статья Google Scholar
Hilton J, Nguyen T, Barbu M, Pei R, Stojanovic M, Lin Q (2012) Полимеразная цепная реакция на основе гранул на микрочипе. Microfluid Nanofluid 13: 749–760
Артикул Google Scholar
Horejsh D, Martini F, Poccia F, Ippolito G, Di Caro A, Capobianchi MR (2005) Молекулярный маяк, анализ на основе шариков для обнаружения нуклеиновых кислот с помощью проточной цитометрии.Нуклеиновые кислоты Res 33: e13
Статья Google Scholar
Howell WM, Jobs M, Gyllensten U, Brookes AJ (1999) Динамическая аллель-специфическая гибридизация. Новый метод оценки однонуклеотидных полиморфизмов. Nat Biotechnol 17: 87–88
Статья Google Scholar
Джобс М., Хауэлл В.М., Стромквист Л., Майр Т., Брукс А.Дж. (2003) DASH-2: гибкое, дешевое и высокопроизводительное генотипирование SNP путем динамической аллель-специфической гибридизации на мембранных массивах.Genome Res 13: 916–924
Статья Google Scholar
Ким С., Мисра А. (2007) Генотипирование SNP: технологии и биомедицинские приложения. Annu Rev Biomed Eng 9: 289–320
Статья Google Scholar
Колпащиков Д.М. (2008) Фермент расщепления ДНК для визуального типирования однонуклеотидного полиморфизма. J Am Chem Soc 130: 2934–2935
Статья Google Scholar
Landegren U, Kaiser R, Sanders J, Hood L (1988) Метод обнаружения генов, опосредованный лигазой.Science 241: 1077–1080
Статья Google Scholar
Lavin MF, Shiloh Y (1997) Генетический дефект при атаксии-телеангиэктазии. Annu Rev Immunol 15: 177–202
Статья Google Scholar
Li Q, Liu Z, Monroe H, Culiat CT (2002) Интегрированная платформа для обнаружения вариантов последовательности ДНК с использованием электрофореза капиллярной матрицы. Электрофорез 23: 1499–1511
Статья Google Scholar
Lim CT, Zhang Y (2007) Микрожидкостные иммуноанализы на основе шариков: новое поколение.Biosens Bioelectron 22: 1197–1204
Статья Google Scholar
Lipsky RH, Mazzanti CM, Rudolph JG, Xu K, Vyas G, Bozak D, Radel MQ, Goldman D (2001) Анализ плавления ДНК для обнаружения однонуклеотидных полиморфизмов. Clin Chem 47: 635–644
Google Scholar
Лямичев В., Маст А.Л., Холл Дж. Г., Прудент Дж. Р., Кайзер М. В., Такова Т., Квятковски Р. В., Сандер Т. Дж., Де Арруда М., Арко Д. А., Нери Б. П., Бровь МАД (1999) Идентификация полиморфизма и количественное определение геномных ДНК путем инвазивного расщепления олигонуклеотидных зондов.Nat Biotechnol 17: 292–296
Статья Google Scholar
Montgomery J, Wittwer CT, Palais R, Zhou LM (2007) Одновременное сканирование мутаций и генотипирование с помощью анализа плавления ДНК с высоким разрешением. Nat Protoc 2: 59–66
Статья Google Scholar
Neo JL, Aw KD, Uttamchandani M (2011) Визуальное генотипирование SNP с использованием асимметричной ПЦР и ферментов расщепленной ДНК.Аналитик 136: 1569–1572
Статья Google Scholar
Ng J, Liu W-T (2006) Миниатюрные платформы для обнаружения однонуклеотидных полиморфизмов. Anal Bioanal Chem 386: 427–434
Статья Google Scholar
Nilsson J, Evander M, Hammarstrom B, Laurell T (2009) Обзор захвата клеток и частиц в микрофлюидных системах. Anal Chim Acta 649: 141–157
Статья Google Scholar
Раменский В., Борк П., Сюняев С. (2002) Несинонимичные SNP человека: сервер и обзор.Nucleic Acids Res 30: 3894–3900
Статья Google Scholar
Riahi R, Mach KE, Mohan R, Liao JC, Wong PK (2011) Молекулярное обнаружение бактериальных патогенов с использованием двухцепочечных ДНК-зондов, усиленных микрочастицами. Anal Chem 83: 6349–6354
Статья Google Scholar
Рогачева М.Б., Фриц К.Л., Рунд Л.А., Поллок С.Б., Бивер Дж. Э., Counter CM, Шук Л.Б. (2007) Характеристика свиного гена ATM: к созданию новой модели атаксии-телеангиэктазии на животных, не являющейся мышиной.Gene 405: 27–35
Статья Google Scholar
Ронаги М., Улен М., Найрен П. (1998) Метод секвенирования на основе пирофосфата в реальном времени. Science 281: 363–365
Статья Google Scholar
Russom A, Haasl S, Brookes AJ, Andersson H, Stemme G (2006) Анализ кривой быстрого плавления на однослойных шариках для высокопроизводительного генотипирования однонуклеотидных полиморфизмов.Anal Chem 78: 2220–2225
Статья Google Scholar
Sandoval N, Platzer M, Rosenthal A, Dörk T, Bendix R, Skawran B, Stuhrmann M, Wegner RD, Sperling K, Banin S, Shiloh Y, Baumer A, Bernthaler U, Sennefelder H, Brohm M, Weber BHF, Schindler D (1999) Характеристика мутаций гена ATM в 66 семьях атаксии-телеангиэктазии. Hum Mol Genet 8: 69–79
Статья Google Scholar
Shendure J, Porreca GJ, Reppas NB, Lin X, McCutcheon JP, Rosenbaum AM, Wang MD, Zhang K, Mitra RD, Church GM (2005) Точное мультиплексное секвенирование полонии эволюционировавшего бактериального генома.Science 309: 1728–1732
Статья Google Scholar
Skelley AM, Kirak O, Suh H, Jaenisch R, Voldman J (2009) Микрожидкостной контроль спаривания и слияния клеток. Nat Methods 6: 147–152
Статья Google Scholar
Sochol RD, Casavant BP, Dueck ME, Lee LP, Lin L (2011) Динамический микрочип на основе шариков для параллельного обнаружения ДНК. J Micromech Microeng 21: 054019
Артикул Google Scholar
Соколов Б.П. (1990) Метод удлинения праймера для обнаружения одиночного нуклеотида в геномной ДНК.Nucleic Acids Res 18: 3671
Статья Google Scholar
Sundberg SO, Wittwer CT, Greer J, Pryor RJ, Elenitoba-Johnson O, Gale BK (2007) Сканирование мутаций ДНК в фазе раствора и генотипирование SNP с помощью анализа плавления нанолитров. Биомедицинские микроустройства 9: 159–166
Статья Google Scholar
Syvanen A-C (2001) Доступ к генетическим вариациям: генотипирование однонуклеотидных полиморфизмов.Nat Rev Genet 2: 930–942
Статья Google Scholar
Такацу К., Йокомаку Т., Курата С., Канагава Т. (2004) Анализ SNP на основе FRET без флуоресцентных зондов. Nucleic Acids Res 32: e156
Статья Google Scholar
Tan W-H, Takeuchi S (2007) Интегрированная микрофлюидная система с улавливанием и выпуском для динамических микрочипов. Proc Natl Acad Sci 104: 1146–1151
Статья Google Scholar
Tong SYC, Giffard PM (2012) Микробиологические приложения анализа плавления с высоким разрешением.J Clin Microbiol 50: 3418–3421
Статья Google Scholar
Тонг А.К., Ли З., Джонс Г.С., Руссо Дж. Дж., Дж. Дж. (2001) Комбинаторные метки переноса энергии флуоресценции для мультиплексных биологических анализов. Nat Biotechnol 19: 756–759
Статья Google Scholar
Tost J, Gut IG (2005) Генотипирование однонуклеотидных полиморфизмов с помощью масс-спектрометрии MALDI в клинических применениях.Clin Biochem 38: 335–350
Статья Google Scholar
Wang Z, Moult J (2001) SNP, структура белка и болезнь. Hum Mutat 17: 263–270
Статья Google Scholar
Уиллер А.Р., Трондсет В.Р., Уилан Р.Дж., Лич А.М., Заре Р.Н., Ляо Й.Х., Фаррелл К., Мангер И.Д., Даридон А. (2003) Микрожидкостное устройство для анализа отдельных ячеек. Anal Chem 75: 3581–3586
Статья Google Scholar
Zhang Y, Park S, Liu K, Tsuan J, Yang S, Wang T-H (2011) Платформа для манипулирования каплями с помощью топографии поверхности для обнаружения биомаркеров и идентификации патогенов.Lab Chip 11: 398–406
Артикул Google Scholar
Общегеномная ассоциация изменений пищевого поведения свиней из-за теплового стресса | Genetics Selection Evolution
Все протоколы для животных соответствовали процедурам, изложенным в Руководстве по уходу и использованию сельскохозяйственных животных в сельскохозяйственных исследованиях и обучении [12], и были одобрены Комитетом по уходу и использованию животных USMARC.
Сбор фенотипических данных
Фенотипические данные были собраны по свиньям в откорме (n = 1648), которые выращивались в U.Центр исследования животных S. Meat с июля 2011 года по март 2016 года. Свиньи помещали в стойло в группах доращивания (n = 7) примерно по 240 свиней в возрасте от 8 до 10 недель. Курганов и свинок смешивали и распределяли по шести загонам, по 39-40 свиней в загоне. Были представлены три линии производителей: дюрок, ландрас и йоркшир, и все самки принадлежали к смешанной популяции ландрас-йоркшир. В среднем в группе отросших братьев было 6,2 полных сибсов и 25,9 отцовских полукровок. Животные были помечены низкочастотной электронной идентификационной меткой при входе в стойло для выращивания.
Загоны были оснащены электронной системой кормления, которая отслеживала поведение при кормлении, как описано Brown-Brandl et al. [13]. Вкратце, у каждого загона была одна кормушка с пятью прорезями, что позволяло есть до пяти животных в любой момент времени. Свиньям был предоставлен неограниченный доступ к рациону из кукурузно-соевого шрота, который был разработан для удовлетворения или превышения потребностей животных в питательных веществах. Каждый слот фидера был снабжен антенной и мультиплексором. Каждые 20 с устройство определяло, какие свиньи находятся у кормушки, а затем записывало количество животных, положение кормушки и время, которые в дальнейшем будут называться «запросами RFID».\ circ {\ text {C}}} \ right) \, {-} \, 14.5} \ right] $$
дней были отнесены к категориям THI на основе максимального значения THI, как указано Брауном-Брандлом и др. [15]. Категории THI включали «Нормальный» (<23,33 ° C), «Предупреждение» (23,33 ° C ≤ × <26,11 ° C), «Опасность» (26,11 ° C ≤ × <28,88 ° C) и «Чрезвычайная ситуация» (≥ 28,88 ° С). Следует отметить, что не все животные попали во все категории THI. Только 949 животных испытали THI выше нормы. Для каждого животного было вычислено общее количество запросов RFID для каждого дня, и среднее количество запросов RFID в день было вычислено для каждой категории THI.Аналогичным образом было вычислено среднее количество запросов RFID в день для каждой породы по комбинации пола для каждой категории THI. Разница между средним числом RFID-запросов животного в определенной категории THI и соответствующим средним значением пола и породы вычислялась и стандартизировалась до среднего значения 0 и стандартного отклонения 1 для каждой категории THI. Различия в поведении кормления между двумя категориями THI (например, Alert-Normal) количественно оценивались путем расчета разницы в стандартизованных пингах RFID между двумя категориями.Следовательно, если животное испытало все четыре категории THI во время фазы завершения, вычислялись четыре стандартизованных значения пищевого поведения THI, которые отражали, как поведение этого животного отклонялось от поведения типичного животного этого типа породы и пола, и были рассчитаны шесть значений, которые указали, как он реагировал на различные температуры по сравнению с его современниками по типу пола и типу породы. Не все животные испытали все четыре категории THI во время завершения роста, и потеря электронных меток привела к разному количеству животных с данными для каждого сравнения.Предполагалось, что животные, которые снизили свою пищевую активность больше, чем их сверстники по типу пола, по мере увеличения категории THI, больше страдали от теплового стресса. Фенотипические корреляции между шестью проанализированными признаками представлены в таблице 1.
Таблица 1 Фенотипические корреляции между шестью проанализированными сравнениями категорий индекса температуры и влажности (THI)Генотипические данные
Образцы хвостов собирали у всех свиней и хранили при -20 ° C. Геномную ДНК экстрагировали с помощью набора для очистки геномной ДНК WIZARD в соответствии с протоколом производителя (Promega Corp., Мэдисон, Висконсин, США). Генотипирование проводилось с использованием трех платформ: чип NeoGen Porcine GGPHD (GeneSeek, Лансинг, США), чип Illumina Porcine SNP60 V2 (Illumina, Inc., Сан-Диего, США) и чип NeoGen GGP-Porcine (GeneSeek, Lansing, США). СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ). Контроль качества включал фильтрацию генотипов, которые имели частоту минорных аллелей ниже 5% и которые не имели уникального положения на карте в сборке генома Sscrofa10.2 [16]. После контроля качества 58 096 однонуклеотидных полиморфизмов (SNP) из чипа GGPHD, 38 598 SNP из чипа Porcine SNP60 V2 и 6882 SNP из чипа GGP-Porcine были сохранены для использования в последующих анализах.Всего было генотипировано 1118 свиней с использованием чипа GGPHD, две свиньи были генотипированы с использованием чипа SNP60 V2, и 34 свиньи были генотипированы с использованием чипа GGP-Porcine. Генотипы животных, генотипированных на чипе Porcine SNP60 V2 и чипе GGP-Porcine, были вменены чипу NeoGen Porcine GGPHD (количество SNP = 58 096) путем вменения родословной с использованием FImpute v2.2 [17].
Полногеномное исследование ассоциации (GWAS)
Каждый из шести признаков (разница между стандартизованным пищевым поведением двух категорий THI) был проанализирован с использованием смешанной линейной модели с полом, породой производителя и современной группой в качестве фиксированных эффектов.Современная группа представляла собой комбинированный эффект группы опороса и загона. Две группы опороса (год – неделя рождения) были представлены в каждой группе, завершившей отрастание, и в стойле было шесть загонов. Хотя фенотипы представляли собой отклонения от пола животного и породы-производителя, порода и пол были включены в качестве фиксированных эффектов для учета стратификации популяции, которая может присутствовать в генотипических данных. Области генома, связанные с каждым признаком, были идентифицированы и количественно оценены с использованием метода отбора переменных Bayes-C и программного обеспечения GenSel [18] на основе следующей модифицированной статистической модели [18]:
$$ {\ mathbf {y}} = {\ mathbf {X}} {\ varvec {\ upbeta}} + {\ mathbf {Zu}} + {\ mathbf {e}}, $$
, где \ ({\ mathbf {y}} \) — вектор признака фенотипы (различия в пищевом поведении между двумя категориями THI), \ ({\ mathbf {X}} \) — матрица случаев фиксированных эффектов (\ ({\ varvec {\ upbeta}} \)), \ ({\ mathbf {Z}} \) представляет собой матрицу генотипов SNP с ненулевыми эффектами (пропорция определяется как 1 — π), которые были подогнаны как случайные эффекты (\ ({\ mathbf {u}} \)), распределенные N (0, \ (\ sigma_ {u} ^ {2} \)), а \ ({\ mathbf {e}} \) — вектор случайных остаточных эффектов, предположительно распределенных нормально N (0, \ (\ sigma _ {{\ mathbf {e}}} ^ {2} \)).
Приоры для генетической и остаточной дисперсии и априорная пропорция SNP, которые, как предполагается, не влияют на признак в рамках итерации цепи Маркова Монте-Карло (MCMC) (π) для каждого признака, были получены путем выполнения Bayes-Cπ с использованием GenSel [18] с той же моделью, что описана выше. Приоры, используемые для анализа Байеса-Cπ, были одинаковыми для всех сравнений категорий THI и составляли 0,98, 0,10 и 0,10 для π, генетической дисперсии и остаточной дисперсии, соответственно. Эти анализы проводились как минимум для 8100 итераций, при этом первые 100 отбрасывались как выгорание.Графики π по итерациям были оценены, чтобы определить, нужны ли дополнительные итерации для получения сходящейся оценки. Результирующие значения π, используемые в анализах Байеса-C, приведены в Таблице 2.
Таблица 2 Апостериорные оценки значений π, полученные из анализа Байеса-Cπ и используемые в анализах Байеса-C для каждой из сравнений категорий индекса температуры и влажности (THI).Для анализа Байеса-C использовалась цепочка из 41 000 итераций, причем первые 1000 циклов отбрасывались как выгорание.Эффекты отбирались каждые 40 итераций для получения апостериорного распределения генетической дисперсии. Геномные области, связанные с каждым признаком, были идентифицированы с использованием геномных окон размером 1 Mb, следуя Wolc et al. [19]. Стандартное отклонение оценок наследуемости на основе маркеров рассчитывалось как стандартное отклонение оценок наследуемости последних 100 выборок.
Функции генов в значимых областях генома
Гены, расположенные в окнах размером 1 Мб, объясняющие более 1.0% геномной дисперсии было получено с использованием аннотации NCBI для Sscrofa10.2 (выпуск 104). Были проанализированы два списка генов. Первый содержал гены, расположенные во всех окнах размером 1 Мб, которые были обнаружены по шести признакам. Во второй список вошли только гены, расположенные в окнах размером 1 Мб, которые объясняют более 3,0% геномной дисперсии хотя бы по одному признаку. Для последнего списка, если два соседних окна превышали 3,0% геномной дисперсии, то в анализ включалась только область размером 1 Mb с наибольшим предполагаемым эффектом.
Система классификации PANTHER (версия 12.0; http://www.pantherdb.org/) [20] использовалась для определения функций генов в этих списках. Обогащающий анализ функции генов был выполнен с использованием реализации PANTHER биномиального теста избыточного представительства [20], который определяет, содержит ли список генов больше генов, участвующих в конкретном пути или функции, чем можно было бы ожидать наугад с поправкой Бонферрони p значение менее 0,05. Значимость терминов генной онтологии (GO) оценивалась с использованием аннотации GO по умолчанию Ensembl Sus scrofa GO в качестве фона для анализа обогащения.
Терморегуляция — обзор | ScienceDirect Topics
Соображения эволюции
Это уже приводит к вопросу о филогенетическом развитии терморегуляции. Чтобы понять эту традицию, может быть полезно взглянуть на некоторые таксоны, которые находятся где-то между экто- и эндотермией. Некоторые насекомые, например, крупные ночные бабочки ( Sphingidae ), пчелы, стрекозы или осы, способны регулировать грудную, а в некоторых случаях и брюшную температуру.Однако эта эндотермия достигается только тогда, когда они активны, они совершают движения крыльями, называемые дрожью, независимо от полета. По крайней мере, бабочки из-за их волосистой чешуи имеют значения теплопроводности, подобные птицам и млекопитающим, и они могут сохранять большие различия между Ta и Tb (некоторые североамериканские бабочки могут летать при температуре тела около 30 ° C при Ta. = 0 ° С). Однако эти мелкие животные не могут достичь постоянной эндотермии, как у позвоночных того же размера, если они не активны днем и ночью (а насекомые — нет).
Среди рыб встречаются более крупные виды в континууме между экто- и эндотермией. Голубой тунец ( Thunnus thymnus ) весом 200–350 кг может выдерживать перепады температур до 20 ° C. У этих рыб, в отличие от «холоднотелых» видов, мы находим большое количество красных (= аэробных) скелетных мышц около ядра тела (вдоль позвоночного столба), а не под кожей. Также высокий BMR и противоточный теплообменник в системе кровообращения являются дополнительными характеристиками этих эндотермических рыб.Помимо красных скелетных мышц, эндотермические рыбы также имеют местные источники тепла в тканях желудка, кишечника и печени. Кроме того (опять же, поддерживаемые retia mirabilia = противоточные теплообменники) в глазах и мозге теплокровных рыб, таких как акулы мако ( Isurus oxyrhynchus ), существует разница температур в окружающей среде> 5 ° C. Однако в головах акул нет тканей, выделяющих тепло, вместо этого теплая кровь из красных мышц живота транспортируется непосредственно в области глаз и мозга.У некоторых костистых рыб (например, Swordfish, Xiphias gladius ), в отличие от акул, глазные мышцы работают как локальные источники тепла, весь комплекс обогревающих мышц, мозга и глаз густо изолирован в жире, а перепады температур достигают 14 ° C может поддерживаться между мозгом и окружающей водой.
Имеются также данные о механизмах контроля физиологической и поведенческой температуры у этих рыб. Также некоторые змеи-питоны и кожистые черепахи ( Dermochelys coriacea ) способны контролировать температуру своего тела.
Таким образом, к вопросу о том, когда и почему могла развиться эндотермия, следует подходить очень широко. Адаптивное значение настоящей эндотермии и эффективной терморегуляции могло заключаться в том, чтобы уменьшить размер тела при постоянной температуре тела. Это позволило бы не только увеличить активность, но и увеличить воспроизводство. Однако эндотермия также требует больших затрат, и поэтому перед ее достижением необходимо выполнить определенные предварительные условия. На биохимическом уровне изменения проницаемости мембран для ионов рассматриваются как необходимые предварительные условия для увеличения скорости метаболизма.На организменном уровне, по крайней мере, в эволюции млекопитающих кажется правдоподобным предположить, что крупные (до 250 кг) рептилии терозавры, предки млекопитающих, благодаря своему большому размеру достигли определенной степени тепловой независимости. и что целый ряд морфологических и физиологических изменений (развитие изолирующей шерсти, повышение эффективности вентиляции за счет развития костного неба и диафрагмы и т. д.) затем позволили перейти от крупных рептилий (с так называемой инерционной гомеотермией, что означает, что они просто были слишком большие, чтобы терять достаточно тепла для того, чтобы быть пойкилотермными) мелким млекопитающим с активной регулирующей эндотермией.
Отображение с высоким разрешением QTL для термостойкости риса с использованием массива SNP 5K | Rice
Begum T, Reuter R, Schöffl F (2013) Сверхэкспрессия AtHsfB4 вызывает специфические эффекты на развитие корней Arabidopsis. Механический Дев 130 (1): 54–60. DOI: 10.1016 / j.mod.2012.05.008
CAS Статья PubMed Google Scholar
Broman KW, Wu H, Sen Ś, Churchill GA (2003) R / qtl: Картирование QTL в экспериментальных скрещиваниях.Биоинформатика 19 (7): 889–890
CAS Статья PubMed Google Scholar
Buu BC, Ha PTT, Tam BP, Nhien TT, Hieu NV, Phuoc NT, Minh LT, Giang LH, Lang NT (2014) Локусы количественных признаков, связанные с толерантностью к жаре у риса ( Oryza sativa L .) Порода растений Биотех 2: 14–24
Статья Google Scholar
Cao L, Zhao J, Zhan X, Li D, He L, Cheng S (2003) Картирование QTL для термостойкости и корреляции между термостойкостью и скоростью фотосинтеза в рисе.Chin J Rice Sci 17: 223–227
CAS Google Scholar
Carriger S, Vallee D (2007) Больше урожая на каплю. Рис сегодня 6: 10–13
Google Scholar
Chang-Lan Z, Ying-Hui X, Chun-Ming W, Ling J, Hu-Qu Z, Jian-Min W. (2005) Отображение QTL для термостойкости на стадии заполнения зерна риса. Rice Sci 12 (1): 33–38
Google Scholar
Chen M, Presting G, Barbazuk WB, Goicoechea JL, Blackmon B, Fang G, Kim H, Frisch D, Yu Y, Sun S, Higingbottom S, Phimphilai J, Phimphilai D, Thurmond S, Gaudette B, Li П., Лю Дж., Хэтфилд Дж., Мэйн Д., Фаррар К., Хендерсон К., Барнетт Л., Коста Р., Уильямс Б., Уолсер С., Аткинс М., Холл С., Будиман М. А., Томкинс Дж. П., Луо М., Бэнкрофт И., Сальсе Дж., Регад Ф., Мохапатра Т., Сингх Н.К., Тьяги А.К., Содерлунд С., Дин Р.А., Винг Р.А. (2002) Интегрированная физическая и генетическая карта генома риса.Растительная клетка 14 (3): 537–545
Статья PubMed PubMed Central Google Scholar
Chen Q, Yu S, Li C, Mou T (2008) Идентификация QTL для устойчивости к теплу на стадии цветения риса. Sci Agric Sin 41: 315–321
CAS Google Scholar
Cheng L, Wang J, Uzokwe V, Meng L, Wang Y, Sun Y, Zhu L, Xu J, Li Z (2012) Генетический анализ устойчивости к холоду на стадии проростков и устойчивости к теплу при цветении риса ( Oryza sativa L.) J Integ Agriculture 11: 359–367
Статья Google Scholar
Дойл Дж., Дойл Дж. Л. (1990) Выделение растительной ДНК из свежей ткани. Фокус 12: 13–15
Google Scholar
Fernandez GCJ (1992) Эффективные критерии отбора для оценки устойчивости растений к стрессу. В: Кус Э.Г. (ред.) Адаптация температуры пищевых культур и водного стресса. Материалы 4-го Международного симпозиума Азиатского центра овощей и исследований и разработок, Шантана, Тайвань, стр. 257–270
Google Scholar
Фишер Р.А., Маурер Р. (1978) Засухоустойчивость сортов яровой пшеницы.I. Ответы на урожайность зерна. Aust J Agric Res 29: 892–912
Google Scholar
Гарг А.К., Ким Дж.К., Оуэнс Т.Г., Ранвала А.П., Чой Ю.Д., Кочиан Л.В., Ву Р.Дж. (2002) Накопление трегалозы в растениях риса обеспечивает высокий уровень толерантности к различным абиотическим стрессам. PNAS 99 (10): 15898–15903. DOI: 10.1073 / pnas.252637799
CAS Статья PubMed PubMed Central Google Scholar
Джорно Ф., Вольтерс-Артс М., Мариани С., Иво Р. (2013) Обеспечение воспроизводства при высоких температурах: реакция на тепловой стресс во время развития пыльника и пыльцы.Растения 2: 489–506. DOI: 10.3390 / растения2030489
Артикул PubMed PubMed Central Google Scholar
Guo J, Wu J, Ji Q, Wang C, Luo L, Yuan Y, Wang Y, Wang J (2008) Полногеномный анализ семейств факторов транскрипции теплового шока у риса и Arabidopsis. Дж. Генет Геномика 35 (2): 105–118. DOI: 10.1016 / S1673-8527 (08) 60016-8
CAS Статья PubMed Google Scholar
IPCC (2007) Резюме для политиков.В: Соломон С., Цинь Д., Мэннинг М., Чен З., Маркиз М., Аверит К. Б., Тигнор М., Миллер Х. Л. (ред.) Изменение климата 2007: основы физической науки. Вклад рабочей группы I в четвертый оценочный доклад межправительственной группы экспертов по изменению климата. Издательство Кембриджского университета, Кембридж, стр. 1–18
Google Scholar
Jagadish SVK, Cairns J, Lafitte R, Wheeler TR, Price AH, Craufurd PQ (2010b) Генетический анализ термостойкости цветков риса.Crop Sci 50: 1633–1641
CAS Статья Google Scholar
Jagadish SVK, Craufurd PQ, Wheeler TR (2007) Высокотемпературный стресс и фертильность колосков в рисе ( Oryza sativa L. ) J Exp Bot 58 (7): 1627–1635
CAS Статья PubMed Google Scholar
Jagadish SVK, Muthurajan R, Oane R, Wheeler TR, Heuer S, Bennett J, Craufurd PQ (2010a) Физиологические и протеомные подходы к решению проблемы термостойкости во время цветения риса ( Oryza sativa L.) J Exp Bot 61: 143–156. DOI: 10.1093 / jxb / erp289
CAS Статья PubMed Google Scholar
Джагадиш С.В., Крауфурд П.К., Уиллер Т.Р. (2008) Фенотипирование родителей картируемых популяций риса ( Oryza sativa L.) на предмет термостойкости во время цветения. Crop Sci 48: 1140–1146
Статья Google Scholar
Kaplan F, Kopka J, Haskell DW, Zhao W., Schiller KC, Gatzke N, Sung DY, Guy CL (2004) Изучение температурно-стрессового метаболома арабидопсиса.Физиология растений 136 (4): 4159–4168
CAS Статья PubMed PubMed Central Google Scholar
Kong D, Li M, Dong Z, Ji H, Li X (2015) Идентификация TaWD40D, белка, содержащего повторы WD40 пшеницы, который связан с устойчивостью растений к абиотическим стрессам. Plant Cell Rep 3: 395–410
Статья Google Scholar
Kumar V, Singh A, Mithra SVA, Krishnamurthy SL, Parida SK, Jain S, Tiwari KK, Kumar P, Rao AR, Sharma SK, Khurana JP, Singh NK, Mohapatra T (2015) Картирование ассоциаций по всему геному солеустойчивости риса ( Oryza sativa L).ДНК Res 22 (2): 133–145. DOI: 10.1093 / dnares / dsu046
CAS Статья PubMed PubMed Central Google Scholar
Li H, Yuan Z, Vizcay-Barrena G, Yang C, Liang W, Zong J, Wilson ZA, Zhang D (2011a) ПОСТОЯННАЯ ТАПЕТАЛЬНАЯ КЛЕТКА1 кодирует белок PHD-finger, который необходим для гибели тапетальных клеток и пыльцы развитие в рисе. Физиология растений 156 (2): 615–630. DOI: 10.1104 / стр.111.175760
CAS Статья PubMed PubMed Central Google Scholar
Li HW, Zang BS, Deng XW, Wang XP (2011b) Сверхэкспрессия гена трегалозо-6-фосфатсинтазы OsTPS1 повышает устойчивость риса к абиотическому стрессу.Planta 234 (5): 1007–1018. DOI: 10.1007 / s00425-011-1458-0
CAS Статья PubMed Google Scholar
Li WL, Liu Y, Douglas CJ (2016) Роль гликозилтрансфераз в образовании примексина стенки пыльцы и формировании паттерна экзин. Plant Physiol. 173; (1): 167–82.
Li N, Zhang DS, Liu HS, Yin CS, Li XX, Liang WQ, Yuan Z, Xu B, Chu HW, Wang J, Wen TQ, Huang H, Luo D, Ma H, Zhang DB (2006 ) Ген задержки дегенерации тапетума риса необходим для деградации тапетума и развития пыльников.Растительная клетка 18: 2999–3014
CAS Статья PubMed PubMed Central Google Scholar
Liu C, Qi X, Zhao Q, Yu J (2013) Характеристика и функциональный анализ специфичного для пыльцы картофеля белка SBgLR, ассоциированного с микротрубочками, в табаке. PLoS One 8 (3): e60543. DOI: 10.1371 / journal.pone.0060543
CAS Статья PubMed PubMed Central Google Scholar
Лю JX, Liao DQ, Oane R, Estenor L, Yang XE, Li ZC, Bennett J (2006) Генетические вариации чувствительности расхождения пыльников к стрессу засухи у риса.Field Crop Res 97: 87–100
Статья Google Scholar
Лобелл Д.Б., Шленкер В., Коста-Робертс Дж. (2011) Климатические тенденции и мировое растениеводство с 1980 года. Наука 333: 616–620
CAS Статья PubMed Google Scholar
Lohmann C, Eggers-Schumacher G, Wunderlich M, Schöffl F (2004) Два разных фактора транскрипции теплового шока регулируют немедленную раннюю экспрессию стрессовых генов у Arabidopsis.Mol Gen Genomics 271 (1): 11–21
CAS Статья Google Scholar
Mansueto L, Fuentes RR, Borja FN, Detras J, Abriol-santos M, ChebotarovD,… Александров N (2017) Обновление базы данных Rice SNP-seek: новые SNP, InDels и запросы. Нуклеиновые кислоты Res 45: D1075-D1081.
Мацуи Т., Омаса К., Хори Т. (1997) Высокая температура, вызванная стерильностью колосков риса японской во время цветения, в зависимости от влажности воздуха и условий скорости ветра.Japan Journal of Crop Science 66: 449–455
Статья Google Scholar
Мацуи Т., Омаса К., Хори Т. (1999) Быстрое набухание пыльцевых зерен в ответ на раскрытие цветочка раскрывает локулу в рисе. Наука о растениеводстве 2 (3): 196–199
Статья Google Scholar
Мацуи Т., Омаса К. (2002) Сорта риса ( Oryza sativa L.), устойчивые к высокой температуре во время цветения: характеристики пыльников.Ann Bot 89 (6): 683–637
Статья PubMed PubMed Central Google Scholar
Meng L, Li H, Zhang L, Wang J (2015) QTL IciMapping: интегрированное программное обеспечение для построения карты генетического сцепления и количественного картирования локусов признаков в популяции с двумя родителями. Урожай J 3: 265–279. DOI: 10.1016 / j.cj.2015.01.001
Артикул Google Scholar
Miranda JA, Avonce N, Suárez R, Thevelein JM, Van Dijck P, Iturriaga G (2007) Бифункциональный фермент TPS-TPP из дрожжей придает трансгенному арабидопсису толерантность к условиям множественного и экстремального абиотического стресса.Planta 226 (6): 1411–1421
CAS Статья PubMed Google Scholar
Мишра А.К., Пураник С., Прасад М. (2012) Структура и регуляторные сети белка WD40 в растениях. J Plant Biochem Biotechnol 21: 32–39
CAS Статья Google Scholar
Мохапатра Т., Робин С., Сарла Н., Шешашай М., Сингх А.К., Сингх К., Сингх Н.К., Амитха Митра С.В., Шарма Р.П. (2014) EMS-индуцированные мутанты сорта риса нагина 22: создание и характеристика.Proc Indian National Science Academy 80: 163–172
Статья Google Scholar
Moon S, Kim SR, Zhao G, Yi J, Yoo Y, Jin P, Lee SW, Jung KH, Zhang D, An G (2013) Rice GLYCOSYLTRANSFERASE1 кодирует гликозилтрансферазу, необходимую для формирования стенки пыльцы. Физиология растений 161 (2): 663–675. DOI: 10.1104 / стр.112.210948
CAS Статья PubMed Google Scholar
Накагава Х., Хори Т., Мацуи Т. (2002) Влияние изменения климата на производство риса и адаптивные технологии.В: Mew TW, Brar DS, Peng S, Dawe D, Hardy B (eds) Наука о рисе: инновации и влияние на средства к существованию Китай: Международный научно-исследовательский институт риса, стр. 635–657
Google Scholar
Nuccio ML, Wu J, Mowers R, Zhou HP, Meghji M, Primavesi LF, Paul MJ, Chen X, Gao Y, Haque E, Basu SS, Lagrimini LM (2015) Экспрессия трегалозо-6-фосфатфосфатазы в початках кукурузы повышает урожайность в условиях полива и засухи. Нат Биотех 33: 862–869.DOI: 10.1038 / NBT.3277
CAS Статья Google Scholar
Pandit A, Rai V, Bal S, Sinha S, Kumar V, Chauhan M, Gautam RK, Singh R, Sharma PC, Singh AK, Gaikwad K, Sharma TR, Mohapatra T, Singh NK (2010) Объединение QTL картирование и профилирование транскриптомов объемных RIL для идентификации функционального полиморфизма генов солеустойчивости у риса ( Oryza sativa L.) Mol Gen Genomics 284 (2): 121–136.DOI: 10.1007 / s00438-010-0551-6
CAS Статья Google Scholar
Poli Y, Basava RK, Panigrahy M, Vinukonda VP, Dokula NR, Voleti SR, Desiraju S, Neelamraju S (2013) Характеристика мутанта риса Nagina22 по термостойкости и картографирование признаков урожайности. Рис 6:36
Артикул PubMed PubMed Central Google Scholar
Prakash C, Mithra SVA, Singh PK, Mohapatra T, Singh NK (2016) Раскрытие молекулярных основ управления окислительным стрессом в засухоустойчивом генотипе риса Nagina22.BMC Genomics 17: 774. DOI: 10.1186 / s12864-016-3131-2
Артикул PubMed PubMed Central Google Scholar
Prasad PVV, Boote KJ, Jr LHA, Sheehy JE, Thomas JMG (2006) Различия между видами, экотипами и сортами в плодовитости колосков и индексе урожая риса в ответ на высокотемпературный стресс. Field Crop Res 95: 398–411
Статья Google Scholar
Qi Y, Liu Q, Zhang L, Mao B, Yan D, Jin Q, He Z (2014) Точное картирование и анализ гена-кандидата нового гена термочувствительной генной мужской стерильности tms9-1 в рисе.Theor Appl Genet 127 (5): 1173–1182. DOI: 10.1007 / s00122-014-2289-8
CAS Статья PubMed Google Scholar
Raman A, Verulkar SB, Mandal N, Variar M, Shukla VD, Dwivedi JL, Singh BN, Singh ON, Swain P, Mall AK, Robin S, Chandrababu R, Jain A, Ram T, Hittalmani S, Haefele S, Piepho HP, Kumar A (2012) Индекс засухи для выбора линий высокоурожайного риса в условиях разной степени стресса от засухи.Рис 5:31. DOI: 10.1186 / 1939-8433-5-31
Артикул PubMed Google Scholar
Сарвар М., Авеси Г.М. (1985) Оценка зародышевой плазмы риса на устойчивость к высоким температурам. Пакистан, журнал J Agric Res 6 (3): 162–164
Google Scholar
Сато К., Инаба К., Тосава М. (1973) Высокотемпературное повреждение созревания растений риса. Влияние высокотемпературной обработки на разных стадиях развития метелки на созревание.Труды Японского общества растениеводства 42: 207–213
CAS Статья Google Scholar
Смертенко А.П., Чанг Х.Й., Вагнер В., Калорити Д., Феник С., Сонобе С., Ллойд С., Хаузер М.Т., Хасси П.Дж. (2004) The Arabidopsis Microtubule-Associated Protein AtMAP65 -1: молекулярный анализ его активность по связыванию микротрубочек. Растительная клетка 16 (8): 2035–2047
CAS Статья PubMed PubMed Central Google Scholar
Szalonek M, Sierpien B, Rymaszewski W, Gieczewska K, Garstka M, Lichocka M, Sass L, Paul K, Vass I, Vankova R, Dobrev P, SzczesnyP MW, Krusiewicz D, Hen-Znigycz , Konopka-Postupolska D (2015) Картофельный аннексин STANN1 способствует устойчивости к засухе и снижает световой стресс у трансгенного Solanum tuberosum L.Растения. PLoS One 10 (7): e0132683. DOI: 10.1371 / journal.pone.0132683
Артикул PubMed PubMed Central Google Scholar
Tazib T, Kobayashi Y, Koyama H, Matsui T (2015) QTL-анализ длины пыльника и растяжения во время цветения как признаков устойчивости риса к экстремальным температурам ( Oryza sativa L.) Euphytica 203: 629– 642
Артикул Google Scholar
Tiwari S, Krishnamurthy SL, Kumar V, Singh B, Rao A, Mithra SVA, Rai V, Singh AK, Singh NK (2016) Отображение QTL для толерантности к соли в рисе ( Oryza sativa L.) посредством массового сегрегантного анализа рекомбинантных инбредных линий с использованием чипа SNP 50K. PLoS One 11 (4): e0153610. DOI: 10.1371 / journal.pone.0153610
Артикул PubMed PubMed Central Google Scholar
Организация Объединенных Наций, Департамент по экономическим и социальным вопросам, Отдел народонаселения (2011 г.) Перспективы народонаселения мира: издание 2010 г., Том I: Комплексные таблицы. ST / ESA / SER.A / 313
Xiao Y, Pan Y, Luo L, Zhang G, Deng H, Dai L, Liu X, Tang W, Chen L, Wang GL (2011a) Локусы количественных признаков, связанные с Завязка семян риса ( Oryza sativa L.) Euphytica 178: 331–338. DOI: 10.1007 / s10681-010-0300-2
Артикул Google Scholar
Xiao YH, Pan Y, Luo LH, Deng HB, Zhang GL, Tang WB, Chen LY (2011b) Локусы количественных признаков, связанные с фертильностью пыльцы в условиях высокотемпературного стресса на стадии цветения риса ( Oryza sativa L ) . Rice Sci 18 (2): 1–7
Google Scholar
Xu L, Tang Y, Gao S, Su S, Hong L, Wang W, Fang Z, Li X, Ma J, Quan W, Sun H, Li X, Wang Y, Liao X, Gao J, Zhang F, Li L, Zhao C (2016) Комплексный анализ семейства генов аннексина в пшенице.BMC Genomics 17: 415. DOI: 10.1186 / s12864-016-2750-у
Артикул PubMed PubMed Central Google Scholar
Ян С., Бискай-Баррена С., Коннер К., Уилсон З.А. (2007) МУЖСКАЯ СТЕРИЛЬНОСТЬ1 требуется для развития тапеталей и биосинтеза пыльцевой стенки. Растительная клетка 19 (11): 3530–3548
CAS Статья PubMed PubMed Central Google Scholar
Ye C, Argayoso MA, Redoña ED, Sierra SL, Laza MA, Dilla CJ, Mo Y, Thomson MJ, Chin J, Delaviña CB, Diaz GQ, Hernandez JE (2012) Картирование QTL для термостойкости на стадии цветения в рисе с использованием маркеров SNP.Порода растений 131 (1): 33–41
CAS Статья Google Scholar
Ye C, Tenorio FA, Argayoso MA, Laza MA, Koh HJ, Redoña ED, Jagadish KS, Gregorio GB (2015) Идентификация и подтверждение локусов количественных признаков, связанных с термостойкостью на стадии цветения в различных популяциях риса. BMC Genet 16:41
Статья PubMed PubMed Central Google Scholar
Zhang C, Zhang F (2015) Многофункциональность белков WD40 в целостности генома и развитии клеточного цикла.J Genomics 3: 40–50
Статья PubMed PubMed Central Google Scholar
Zhang G, Chen L, Xiao G, Xiao Y, Chen X, Zhang S (2009) Объемный сегрегантный анализ для определения QTL, связанного с термостойкостью риса, с использованием маркеров SSR. Сельское хозяйство Китая 8: 482–487
CAS Статья Google Scholar
Zhang T, Yang L, Jiang K, Huang M, Sun Q, Chen W, Zheng J (2008) Отображение QTL для термостойкости периода кисточки риса.Mol Plant Breed 6: 867–873
CAS Google Scholar
Zhao L, Lei J, Huang Y, Zhu S, Chen H, Huang R, Peng Z, Tu Q, Shen X, Yan S (2016) Картирование локусов количественных признаков термостойкости при цветении риса с использованием хромосомного сегмента строки подстановки. Breeding Science Предварительный просмотр . DOI: 10.1270 / jsbbs.15084.
Zhu J, Wu X, Yuan S, Qian D, Nan Q, An L, Xiang Y (2014) Annexin5 играет жизненно важную роль в развитии пыльцы Arabidopsis через Ca 2+ -зависимый мембранный перенос.PLoS One 9 (7): e102407
Артикул PubMed PubMed Central Google Scholar
Анализ плавления с высоким разрешением | Обзор HRM
Ссылки >> HRM
HRM технологииВведение в HRM
Плавление с высоким разрешением (HRM) — это однородный, очень мощный метод для генотипирования SNP, сканирования мутаций и сканирования последовательностей в образцах ДНК. Благодаря появлению в последнее время улучшенных красителей, связывающих двухцепочечную ДНК (дцДНК), и оборудования для ПЦР нового поколения в реальном времени, анализ плавления с высоким разрешением основан на методах кривой плавления (диссоциации) ПЦР.
Технология HRM характеризует образцы нуклеиновых кислот на основе их поведения диссоциации и обнаруживает небольшие различия последовательностей в последовательностях, амплифицированных ПЦР, просто путем прямого плавления. Образцы также различают по длине последовательности, содержанию GC и комплементарности цепи. Эти различия обнаруживаются с помощью специальных красителей для ДНК, высококлассного оборудования и сложного программного обеспечения для анализа.
Механизм
Анализ плавления с высоким разрешением (HRM) — это метод после ПЦР.Интересующий участок в последовательности ДНК сначала амплифицируется с помощью полимеразной цепной реакции. Во время этого процесса в реакцию добавляются специальные насыщающие красители, которые флуоресцируют только в присутствии двухцепочечной ДНК. Такие красители известны как интеркалирующие красители.
Во время ПЦР амплифицированная интересующая область известна как ампликон. По мере увеличения концентрации ампликона в реакционной пробирке флуоресценция, проявляемая двухцепочечным амплифицированным продуктом, также увеличивается.
После процесса ПЦР начинается анализ HRM. В этом процессе ДНК ампликона постепенно нагревается от примерно 50 ° C до примерно 95 ° C. По мере повышения температуры в какой-то момент достигается температура плавления ампликона, и образец ДНК денатурируется, а двухцепочечная ДНК расплавляется. Благодаря этому флуоресценция гаснет.
Это связано с тем, что в отсутствие двухцепочечной ДНК интеркалирующим красителям не с чем связываться, и они флуоресцируют только на низком уровне.Это наблюдение нанесено на график, показывающий уровень флуоресценции в зависимости от температуры, с получением кривой плавления. Даже одно изменение основания в последовательности ДНК образца вызывает различия в кривой HRM. Поскольку разные генетические последовательности плавятся с несколько разной скоростью, их можно просматривать, сравнивать и обнаруживать с помощью этих кривых.
Кривые плавления, полученные после анализа плавления с высоким разрешением, обычно строятся с флуоресценцией по оси Y и температурой по оси X. Они похожи на графики амплификации ПЦР в реальном времени, но с заменой номера цикла на температуру.
Рисунок: Кривая плавления ДНК для анализа плавления с высоким разрешением
Интеркалирующие красители для HRM-анализа
Насыщающие флуоресцентные красители используются для контроля денатурации ампликонов ДНК в образце с целью проведения HRM-анализа. Некоторые из имеющихся в продаже красителей:
LC Зеленый
Красители LC Green специально разработаны для анализа плавления с высоким разрешением для обнаружения вариантов последовательностей ДНК.Добавление этих красителей увеличивает температуру плавления ДНК на 1-3 ° C и может потребовать корректировки параметров цикла.Краситель SYTO
Красители SYTO представляют собой проницаемые для клеток красители нуклеиновых кислот, которые демонстрируют значительное усиление флуоресценции, когда они связываются с двухцепочечными последовательностями нуклеиновых кислот. Эти красители коммерчески доступны как синие, зеленые, оранжевые или красные флуоресцентные красители.EvaGreen®
Краситель EvaGreen® — это зеленый флуоресцентный краситель на основе нуклеиновых кислот, широко используемый для анализа плавления с высоким разрешением.Краситель сам по себе не является флуоресцентным, но становится очень флуоресцентным при связывании с двухцепочечной ДНК. Он не является мутагенным, нецитотоксическим и полностью непроницаем для клеточных мембран.- Chromofy
Chromofy — мономерный асимметричный цианиновый краситель. Когда chromofy связывается с двухцепочечной ДНК, она показывает очень сильное усиление флуоресценции. Chromofy можно использовать для анализа HRM для анализа генотипирования SNP и метилирования.
Преимущества HRM
Рентабельность
Анализ расплава ДНК с высоким разрешением экономически эффективен, чем другие технологии генотипирования, такие как секвенирование и типирование SNP Taqman.Это делает его идеальным для крупномасштабных проектов генотипирования.Простой и быстрый рабочий процесс
После выполнения ПЦР-амплификации целевой последовательности никаких дополнительных инструментов для HRM-анализа не требуется.Быстрый и мощный
HRM Analysis — это быстрый и мощный инструмент, благодаря которому он может точно генотипировать огромное количество образцов за короткое время с высоким уровнем точности.Низкий расход реагентов
Для анализа плавления с высоким разрешением требуется только объем реакции ПЦР для анализа каждого образца, что исключает необходимость использования растворителей для ВЭЖХ или гелей для денатурирующего градиентного гель-электрофореза (DGGE).
Приложения HRM
SNP Генотипирование
Картирование ДНК
Сканирование мутаций
Идентификация видов
Проверка зиготности
Анализ метилирования ДНК
Отпечаток ДНК
Beacon Designer ™ разрабатывает высокоспецифичные и эффективные праймеры HRM для помощи в генотипировании SNP.